1. Introduction
Virus diseases are considered potential culprits for the regional population decreases in honey bees [
1] and wild pollinators [
1,
2] despite the absence of a decreasing trend in the global honey bee population [
3]. In recent years, the over-winter survival rate of honey bees in the US has remained consistently low, and this decline has been attributed to viruses [
4,
5]. Despite the potential threats posed by viruses to honey bee health, our limited understanding of virus biology and pathogenesis presents a significant obstacle to the development of effective disease treatments and management strategies.
Most honey bee-infecting viruses documented thus far belong to the
Picornavales order, characterized as single-stranded positive-sense RNA (+ssRNA) viruses. These viruses exhibit similarities in terms of viral capsid, genome structure, and protein translation with picornaviruses [
6], which are notorious for causing severe diseases in humans, such as polio and enterovirus diseases, as well as in animals, such as foot and mouth disease in swine. Extensive research on these picornaviruses has provided in-depth insights into virus–host interactions [
7,
8,
9], which hold the potential for developing pharmaceutical solutions [
10,
11,
12]. However, applying the findings directly from picornavirus studies to bee-infecting RNA viruses is challenging without conducting de novo studies involving the genetic manipulation of the virus under controlled conditions. Consequently, our understanding of viral gene function and detailed virus–host interactions in honey bee-infecting RNA viruses is still relatively limited compared with the knowledge amassed for picornaviruses.
Creating infectious clones of targeted bee-infecting viruses represents the initial step in manipulating virus genomes. To date, numerous clones of bee-infecting viruses have been synthesized [
13,
14,
15,
16,
17], and the use of these clones in application studies is increasingly prevalent [
18,
19]. As these viruses possess a single-stranded positive-sense RNA genome that mimics mRNA, the synthesized viral genome DNA templates are commonly referred to as cDNA templates. The process of editing the viral sequence to investigate gene functions is often referred to as reversed genetics [
20].
However, implementing the standard method for producing infectious cloned viruses can present significant technical challenges [
21]. Firstly, the need to produce full-length viral RNA in 8–10 kb sizes, which is considerably larger than the typical sizes required for gene expression or RNA interference experiments, poses a major obstacle for researchers. Moreover, RNA is susceptible to degradation, and transcribed RNA requires ultra-low temperatures during shipping. The in vitro transcription process may also be challenging to replicate in different laboratory settings, particularly those not specialized in viral or molecular biology. Furthermore, the inherent difficulties associated with replicating studies are exacerbated by these challenges. In our own research, we encountered instances where the full-length viral genome RNA of our cloned SBV [
14] degraded into smaller sizes during the repetition of in vitro transcription. While quality control measures can help address this issue, they require significant time and labor resources.
Secondly, the viability of produced infectious clones may gradually diminish during storage and shipment. Currently, there is no viability assay available for bee-infecting RNA viruses, making it impossible to conduct titration assays before each inoculation. This limitation may lead to difficulties in comparing the results published using somewhat different settings. These obstacles further compound the challenges faced by laboratories involved in interdisciplinary and geographically distant collaborations when working with infectious clones.
Lastly, the requirement of inoculating viral RNA into live bees to produce infectious clones can be problematic due to the scarcity of virus-free honey bees. In addition to contaminations of the infectious clones with other viruses that have infected the testing hosts, mixed infections of wild-type and cloned viruses of the same virus species can easily deviate from the conditions necessary for generating robust and repeatable results. In our previous study of the SBV clone, we encountered difficulties due to the lack of SBV-free bees during the repeating trials [
14].
The utilization of a cDNA template with self-initiating transcription capability for viral genome transcription within cultured cells offers a potential solution to the challenges associated with the use of infectious virus clones [
22]. Distributing the infectious clones in DNA plasmid status would be much easier than the RNA or the purified virions. Using a DNA plasmid directly to synthesize virus clones can largely reduce difficulties, cost, and labor. Additionally, plasmids provide greater convenience for sequence editing [
22]. These features would streamline the reversed genetic studies and collaborations that apply the infectious clones in various honey bee research endeavors.
In this article, we present the first attempt to create a cDNA template that can synthesize an infectious clone of a honey bee-infecting RNA virus. We used the AcMNPV immediate-early protein (IE1) promoter to generate a Chinese variant SBV clone with an EGFP expression [
14]. In our previous study, this clone demonstrated infectivity and caused overt infections. AcMNPV is the most commonly used baculovirus vector in insect cell lines, and the IE1 promoter can initiate the transcription within various host cells without assistance from other viral proteins [
23]. In addition to testing the IE1-SBV clone in
A. mellifera and
A. cerana pupae, we also evaluated its potential to infect an immortal lepidopteran cell line derived from
Spodoptera frugiperda, known as Sf9, as an alternative host (not a natural host) for the cloned virus.
2. Materials and Methods
2.1. Ethics Statement
No commercial interests were generated from or involved in this study. All materials used in this research were properly stored and handled without being released into the field. The pGL3-IE1 plasmid was provided by Addgene (addgene.org), a non-profit repository that facilitates the sharing and distribution of plasmids among scientists. The pGL3-IE1 plasmid was obtained through their distributor in China, Beijing Zhongyuan, Ltd.
2.2. Plasmid Construction for Synthesis of the SBV Clone Induced by IE1 Promoter
We selected the immediate-early protein (IE1) promoter derived from the baculovirus Autographa californica multiple nucleopolyhedrovirus (AcMNPV) to drive the synthesis of the cloned virus in host cells. The IE1 promoter sequence has been extensively used for expressing various proteins in cell lines and live insects [
24]. For our study, we utilized the pGL3-IE1 plasmid, which has been previously employed in live insects [
25] and was generously provided by Dr. Zach Adelman (Addgene plasmid #52894). The pGL3-IE1 plasmid contains an origin of replication (pBR322) and antibiotic-resistance that matches the vector used in our previous study [
14] for cloning the SBV genome, indicating feasibility of transferring the cloned SBV genome into the pGL3-IE1 plasmid.
The sacbrood virus clone (GenBank accession number MN528599) used in this study was obtained from
Apis cerana larvae collected in Fuzhou, China. Although this isolate shares genetic similarity with Chinese variants, also known as Chinese SBV (CSBV), it exhibits only slight genetic differences compared with the typical SBV genome found in
A. mellifera populations in the US and Europe. Moreover, this variant is prevalent in honey bee populations across East and Southeast Asia. In a previous study [
26], I referred to this variant as AcSBV, but it is possible that
A. mellifera populations may also be infected by this variant in the field [
27,
28]. Given the taxonomical uncertainty associated with this variant and that the main focus of this study is not the phylogenetic analysis of SBV variants, we consistently use the abbreviation “SBV” throughout the article.
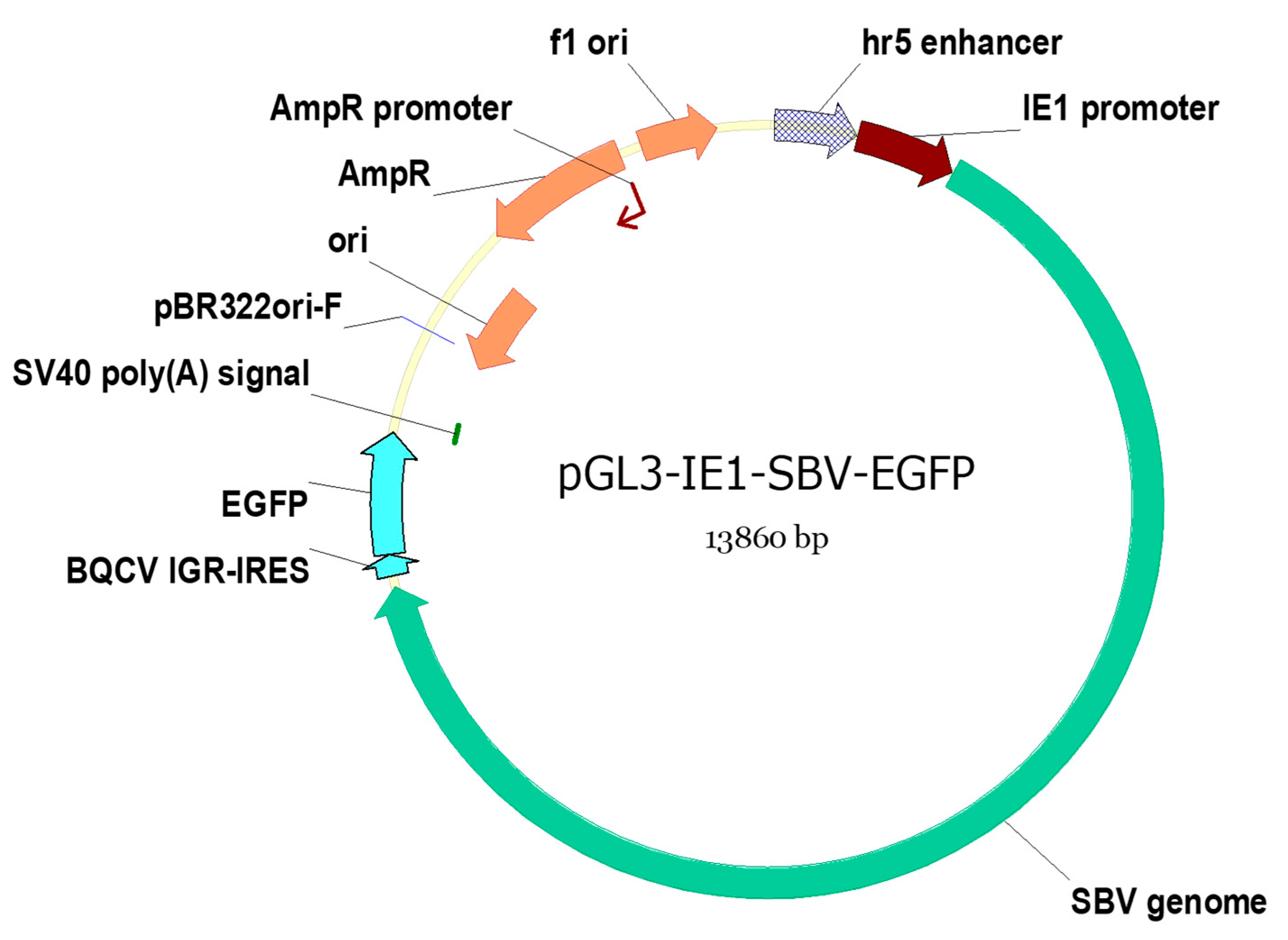
The SBV genome and pGL3-IE1 plasmid were amplified using PCR with the primers designed to replace the luciferase gene region on pGL3-IE1 with the SBV clone. This SBV clone utilized enhanced green fluorescence protein (EGFP) expression tag inserted within the 3′ untranslated region (UTR) [
14]. The translation of EGFP is driven by a separate internal ribosome entry site (IRES) cloned from Black queen cell virus (BQCV IGR-IRES, as shown in
Figure 1). The design of the EGFP-expressing tag ensures its dissociation from the cloned SBV open reading frame (ORF) as previous attempts to mutate sequences within the ORF were unsuccessful. Previously, this identical clone has been demonstrated to induce overt infections in
A. cerana larvae, and its estimated infectious dose was determined in our previous study. [
14].
The SBV clone PCR was conducted using the primer set T7-SBV5′ and SV40-EGFP-r. The pGL3-IE1 plasmid was also PCR amplified using the primer set T7-IE1-R and SV40F. These primers (listed in
Table 1) included an additional overlapping region at 5′ end to facilitate Gibson assembly. Phanta Max Super-Fidelity DNA Polymerase (Vazyme, Nanjing, China) was used for the PCR, and the PCR settings and recipes followed the annotations in
Table 1 and the manufacture’s suggestions, which were identical to those used in our previous study [
14]. The poly(A) region of the original clone was removed since the designated pGL3-IE1 plasmid has a simian virus 40 (SV40) poly(A) signal sequence that adds poly(A) region to the transcribed RNA (
Figure 1).
The pGL3-IE1-SBV-EGFP plasmid (referred to as IE1-SBV) was constructed using the Gibson assembly method [
30] and the pEASY-Uni Seamless Cloning kit from Transgene. The map of the assembled plasmid is shown in
Figure 1. The assembled plasmid was transfected into Stbl3
E. coli competent cells (Shanghai Weidi Biotechnology) using the suggested method, with the incubation temperature set at 30 °C. The transfected
E. coli cells were cultured on LB plates with ampicillin at 30 °C for 20 h. Selected bacterial colonies were propagated in 5 mL LB medium with ampicillin at 30 °C (on an orbital shaker at 200 rpm) for 20 h. Three milliliters of the cultured
E. coli solution was used for plasmid extraction using E.Z.N.A. Plasmid DNA Mini kit (Omega), while the remainder was mixed with glycerol (approximately 20% by volume), thoroughly mixed, and stored at −80 °C for further propagation. The extracted plasmids were linearized using EcoRI and their sizes were determined through gel electrophoresis. Plasmids of the expected size were subjected to Sanger sequencing using ABI 3730XL (performed by BioSune, Shanghai, China). The sequencings were conducted using known primers near the SBV genome termini [
14] and the known primers were listed on pGL3-IE1 map (Addgene plasmid #52894) to confirm the correctness of the assembly. The
E. coli culture containing the correct plasmid was further propagated by transferring 100 μL of the culture stored at −80 °C into 200 mL fresh LB with ampicillin. The culture was incubated at 30 °C (on an orbital shaker at 200 rpm) for 20 h. The plasmid was isolated using FastPure Endo-free Plasmid Maxi kit (Vazyme).
This plasmid is modified from the pGL3-IE1 plasmid [
25] that contains IE1 promoter with hr5 enhancer to express the SBV clone, and the SV40 poly(A) signal adds poly(A) region to the transcribed SBV clone. The EGFP expressing tag is induced by a separated IGR-IRES cloned from BQCV, and it is independent of the clone infectivity [
14]. Elements on the figures are as follows: f1 ori (f1 bacteriophage origin of replication); pBR322ori (origin of replication from pBR322 plasmid, resulting in approximately 15–20 copies per cell); AmpR: ampicillin resistance; SV40 poly(A) signal (Simian virus 40-derived signal that facilitates polyadenylation of transcribed RNA); and hr5 enhancer and IE1 promoter (hr5, homologous region 5, fused with the immediate-early gene (IE1) promoter, from Autographa californica multiple nucleopolyhedrovirus, AcMNPV. IE1 promoter enables transcription initiation without the requirement of other viral proteins).
2.3. Transfection of the IE1-SBV Plasmid in Sf9 Cells
The conventional cationic liposome method was employed to deliver the IE1-SBV plasmid DNA into cultured cells and honey bee pupae. Since a virus-free immortal cell line of honey bees was not yet widely available at the time, the commonly available insect cell line originating from
Spodoptera frugiperda, Sf9 (Procell Life, Wuhan, China), was utilized as the in vitro host cells for IE1-SBV in serum-free culture medium, Sim SF medium (Sino-biological, Beijing, China). Lipofectamine 2000 (Invitrogen, Waltham, MA, USA) was used as the liposome reagent for the transfection. Lipofectamine 2000 has been widely used in studies, including live insect injections for RNAi trials [
31].
The Sf9 cells were transfected according to the recommended protocol for adherent cells using Lipofectamine 2000. Half-density cells (4 × 105) were seeded on each well of 6-well plates and transfected with 4000 ng of the IE1-SBV plasmid or pGL3-IE1 plasmid per well. The plasmid DNA was diluted into a volume of 250 μL using Sim SF medium (Sino-biological, China), with 4000 ng of plasmid DNA. This diluted plasmid DNA was then mixed with 250 μL Lipofectamine solution, prepared by combining 240 μL Sim SF with 10 μL Lipofectamine 2000. The mixture was incubated at room temperature for 20 min. Subsequently, the mixture was added to a well of 6-well plates containing seeded Sf9 cells, along with 1.5 mL Sim SF medium. Each plasmid tested underwent three repeats, with three wells allocated for each repeat.
2.4. Plasmid Injection in Honey Bee Pupae
To assess the ability of IE1-SBV to synthesize the clone in honey bees, we conducted injections of IE1-SBV and pGL3-IE1 (empty vector) plasmids using Lipofectamine 2000 (Invirtrogen) into honey bee pupae. A modified injection protocol was developed based on preliminary trials using Lipofectamine 2000 mixed with plasmid solutions at different concentrations. Initial tests with various concentrations of the IE1-SBV plasmid were performed in A. mellifera to determine the optimal amount for injection. Following the manufacturer’s manual, which suggested a ratio of 2–4 μL Lipofectamine 2000 per 1000 ng of DNA, we decided to use 4 μL Lipofectamine 2000 mixed with 4 μL of DNA plasmids, totaling 8 μL. This volume was selected to be 20% lower than the maximum capacity for pupa injection to enhance feasibility. To determine the maximum capacity, we injected PBS with blue ink into 24 pupae and observed any potential leakage and distribution of the injection within the bee pupae.
In the first trial, three groups of eight A. mellifera pupae were injected with 750, 1000, and 1200 ng of the IE1-SBV plasmid DNA. Among the injected pupae, only those in the groups receiving 1000 and 1200 ng of the IE1-SBV plasmid exhibited detectable virus infection. However, the infection rate was low, with only one and two out of the eight injected pupae showing infection in the RT-qPCR exam. Therefore, we continued with pupae injections and increased the amount of injected plasmid to 1800 ng in a subsequent trial using A. mellifera pupae in December 2021.
The injection protocol involved concentrating the plasmid using a 10 kDa centrifugal filter (0.5 mL, Millipore) and diluting it in Sim SF medium (Sino-biological, China) to the desired concentrations before mixing it with Lipofectamine 2000. The mixture was prepared in bulk for 20 pupae (160 μL in total) and incubated for at least 20 min at room temperature. Each pupa was then injected with 8 μL of the mixture, which contained 1800 ng plasmid DNA and 4 μL Lipofectamine 2000. Pupae in the white- to pink-eye stage (within 2–3 days after pupation) were carefully extracted from capped cells using a rubber air blower and soft entomology forceps. The pupae were transferred to a Petri dish in a 35 °C growth chamber with 70% relative humidity. For each injection, 12–20 pupae were taken out of the growth chamber, and a LeTkingok syringe propeller with a 1-mL disposable syringe and a 30 G (for the trial injecting 1800 ng plasmid) or 32 G (for the trial injecting 2500 ng) needle was used to inject the 8 μL plasmid mixture into each pupa over a period of 6 s. The injected pupa was then held still with the needle for an additional 10 s to allow the injected liquid to distribute evenly within the pupa and minimize post-injection leakage. Pupae that did not exhibit significant leakage after injection were transferred to a 96-well cell culture plate and returned to the 35 °C growth chamber for a 5-day incubation period. After incubation, each pupa was transferred into an Eppendorf tube containing 250 μL RNA keeper (Vazyme) and stored according to the manufacturer’s suggestions until RNA extraction.
In December 2022, we identified a few A. cerana colonies that had no detectable SBV infections. We conducted a similar trial by injecting different concentrations of the IE1-SBV and pGL3-IE1 plasmids (1250, 2500, 5000 ng) into A. cerana pupae harvested from two different A. cerana colonies. Each group had 16 pupae injected, and 12 pupae were selected for RT-qPCR analysis using the TaqMan method for SBV quantification. The results showed that 2500 ng resulted in the highest infection ratio among the tested concentrations. Subsequently, we conducted a parallel trial using A. mellifera pupae and A. cerana pupae from another colony. Twelve pupae were injected with 2500 ng of the IE1-SBV or pGL3-IE1 plasmid, and eight of the twelve pupae were randomly selected for the RT-qPCR examination.
2.5. Cell Preparation and Staining for Confocal Microscopy Analysis
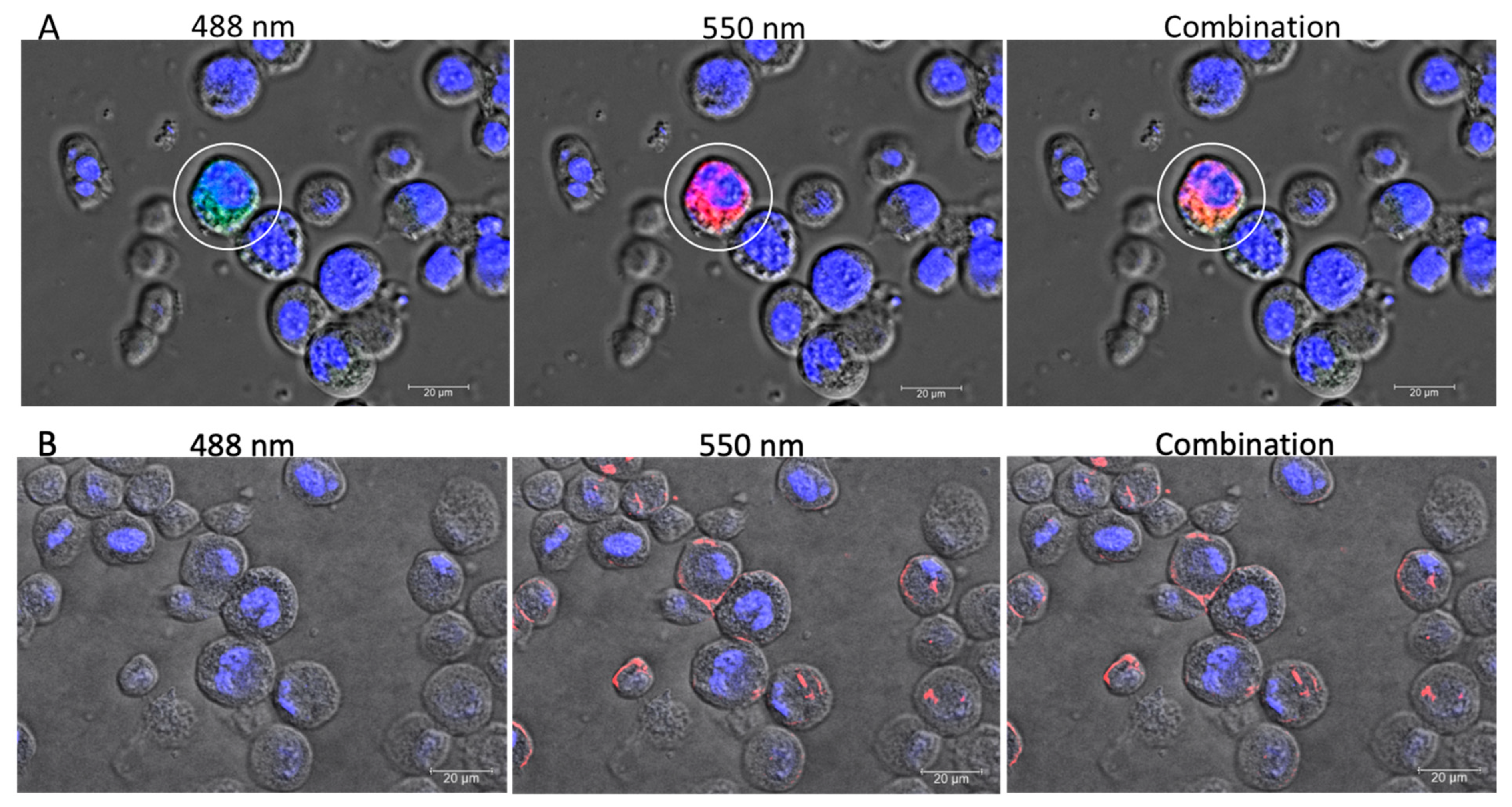
To facilitate handling and observation under a confocal microscope, the Sf9 cells were seeded on cover glasses (WHB scientific, Shanghai, China) placed within 6-well plates before the transfection. The same transfection protocol using Lipofectamine 2000, as described earlier for the 6-well plates, was employed for transfecting IE1-SBV or pGL3-IE1 (original plasmid without SBV genome) plasmids. After 48 h incubation, the cover glasses were removed from the medium and rinsed twice in PBS. They were then transferred into 4% paraformaldehyde in PBS as a fixative for 15 min. Subsequently, the cover glasses were rinsed twice in PBS to remove the fixative.
To reveal nuclei, the cover glasses were stained in DAPI solution (a fluorescent DNA stain; Solar-bio, Beijing, China) followed by staining with Dil stain (a lipophilic fluorescent stain for membrane structures; Solar-bio, Beijing, China). The cover glasses were washed twice in PBS and then embedded in an anti-fade mounting medium (glycerol-based, Solar-bio, Beijing, China). Preliminary observations indicated that cells expressing EGFP fluorescence, the expressing tag of IE1-SBV plasmid, appeared to have over-developed membrane structures under visible light observation. To differentiate the membrane structures in transfected and normal cells, Dil stain was added. A low concentration of Dil stain (PBS with 0.02% of the stock solution, 1 mg/mL in DMSO) was used to prevent over-exposures.
The cells were observed using a Leica SP8 confocal microscope.
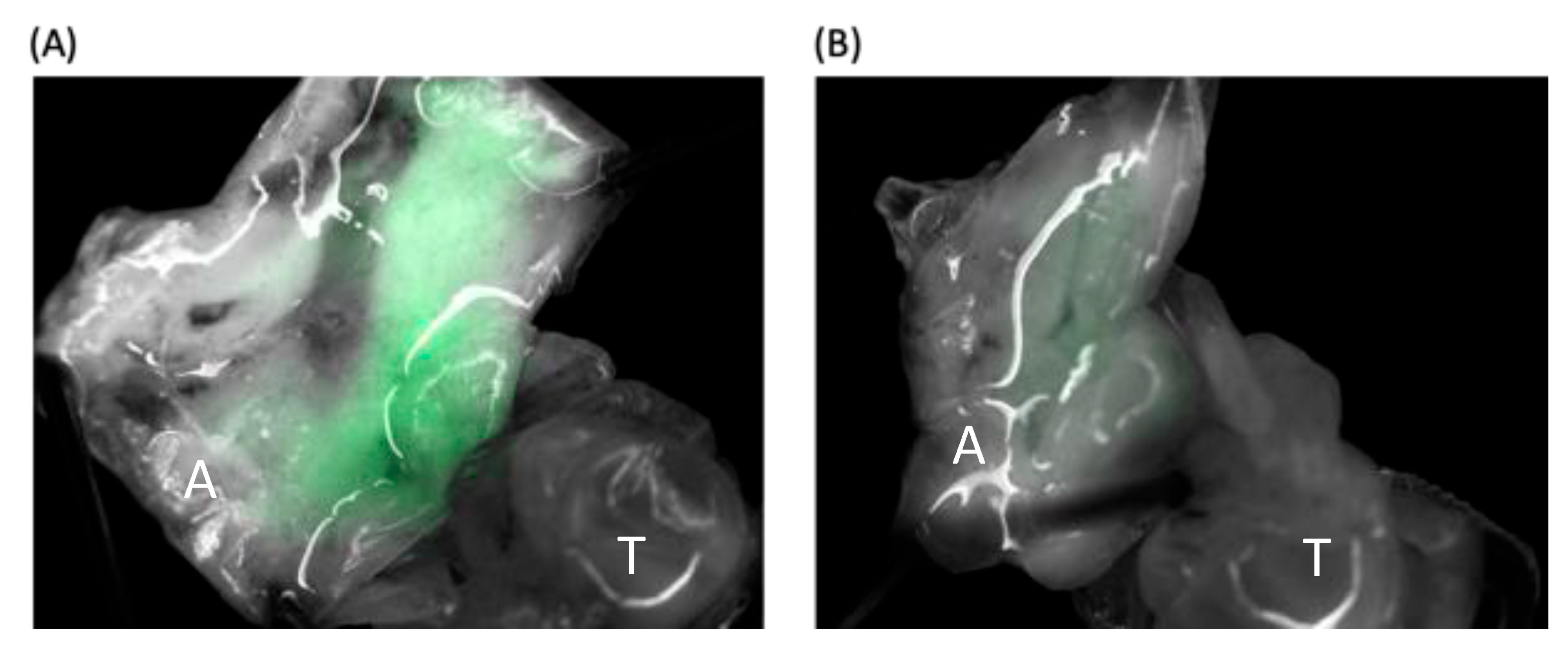
2.6. Plasmid-Injected Pupa Observations under a Stereo Fluorescence Microscope
To reveal the EGFP expression in honey bee pupae, the pupae were observed under a fluorescence stereo microscope (Leica M205 FA). The abdomens were dissected using a surgical scalpel, and subsequently secured in place using insect pins to reveal the pupal tissues under the Leica M205 FA microscope. After the observation, the pupa was transferred into tubes with RNA keeper (Vazyme). Only the last repetition of the IE1-SBV and pGL3-IE1 plasmid (1800 ng)-injected A. mellifera pupae were observed under the fluorescence stereo microscope.
2.7. RNA Extraction and Reverse Transcription for SBV Clone Infection Quantification
The positive strand of the SBV clone was quantified using reverse transcription quantitative polymerase chain reaction (RT-qPCR) to assess the infection intensities. For pupae, total RNA was extracted individually by homogenizing each pupa in 250 μL RNA keeper (R501, Vazyme) and subjecting 30 μL homogenate to RNA extraction using LabServ Universal RNA kit (Thermo-Fisher, Waltham, MA, USA) with a KingFisher (Thermo-Fisher) automatic manipulator, following the manufacturer’s suggestions. The extracted RNA was eluted into 30 μL RNase-free water and then reverse transcribed.
For Sf9 cells, the culture medium was removed, and the cells were rinsed with PBS twice to remove most of the medium residues. Plasmid residues were eliminated by the addition of DNaseI (Invitrogen) according to the protocol provided in the manual. After removing the liquid from the well, 1 mL of the lysis buffer from the LabServ kit (Thermo-Fisher) was added to each well of the 6-well plate, and the cells were scraped from the surface using a rubber scraper. The cell–lysate mixture was transferred into 1.5 mL Eppendorf tubes and then processed using the manufacturer’s suggested methods to extract total RNA.
We employed a two-step RT-qPCR protocol with absolute quantification to determine the SBV genome copy numbers. The same amount of RNA from Sf9 cell lysates and honey bee pupa homogenates were used for cDNA synthesis. RNA samples were quantified using Nanodrop (Thermo-Fisher) and 1000 ng of each RNA sample was used for cDNA synthesis. The cDNA was all synthesized using HiScript II Q Select kit with gDNA remover (R232 kit, Vazyme) following the manufacturer’s protocol. The qPCR was performed using the universal SBV TaqMan method [
29] (primers and probe were listed in
Table 1) with AceQ qPCR Probe Master Mix (Vazyme). The IE1-SBV plasmid DNA was used as the standards in absolute quantification PCR. The plasmid DNA concentration was measured using a Nanodrop (Thermo-Fisher) and copy numbers were estimated based on the molecular weight of the plasmid sequence. A standard curve was generated by serial dilution (10-fold dilution of seven concentrations) in each qPCR. The SBV copy numbers were estimated in CFX manager 3.1 (Biorad, Shanghai, China).
Upon analyzing our preliminary results, we identified the potential for IE1-SBV plasmid DNA residues within our extracted RNA to influence quantification. To mitigate this risk, we implemented a quality control process for all synthesized cDNA by employing a primer set tailored to the IE1 promoter region (
Table 1). As this region is not transcribed into RNA, any presence of IE1-SBV plasmids within the cDNA renders the RNA samples and results unusable. The quality control was conducted using qPCR with SYBR green methodology, employing ChemQ Universal SYBR qPCR Master Mix (Vazyme) and 400 pM primers in a three-step program, with 95 °C for 10 s and 60 °C for 10 s, followed by 72 °C for 20 s, for 40 cycles.
2.8. Tagged RT-PCR to Assess Negative Strand Presence
Tagged RT-PCR was employed to investigate the presence of the negative strand in RNA samples isolated from transfected Sf9 cells and injected honey bee pupae. This evaluation allowed us to assess whether the expressed viral genome RNA was functional and capable of replications. We included two RNA samples of each group from the pupae injection that exhibited positive results and high SBV copy counts in the RT-qPCR examination for the detection of the negative strand. Additionally, all RNA samples from Sf9 cells transfected with either the IE1-SBV or pGL3-IE1 plasmid were included in the detection process. Due to the low copy numbers obtained in the quantification PCR results, a semi-nested PCR approach utilizing two sequential PCRs was applied to enhance the sensitivity and specificity of the assay. Tagged primer (
Table 1) was used in cDNA synthesis using a HiScript II kit with gDNA remover (R212 kit, Vazyme) following the manufacturer’s suggested protocol, except for doubling the DNase treatment time. The cDNA samples were subjected to the first PCR using the tag primer and primer R1 (
Table 1), whilst 0.5 μL cDNA was used in a 25 μL reaction using Phanta Max Super-Fidelity DNA Polymerase (Vazyme). The PCR products were diluted 10 times, and then 1 μL of the dilutions were subjected to the second PCR using the same tag primer and R2 (
Table 1). A negative control, identical in all aspects except for the absence of reverse transcriptase, was conducted for each RNA sample to examine the presence of any plasmid DNA residue or unspecific detection. The PCR products were visualized using 1% agarose gel electrophoresis.
4. Discussion
The SBV clones were synthesized using DNA plasmids with the IE1 promoter in Sf9 cell lines and honey bee pupae. In our previous study, we demonstrated that this identical clone caused overt infection and allowed for the estimation of infection doses in
A. cerana larvae [
14]. Although Sf9 cells and the pupal stage of
A. mellifera and
A. cerana are not the typical hosts for overt SBV infection, they offer convenience for laboratory manipulation and maintenance. Intriguingly, our previous trial involved the injection of transcribed viral genome RNA from the identical clone into
A. mellifera pupae [
14] and yielded infection ratios comparable to those observed in the injection of the IE1-SBV plasmid in
A. mellifera pupae. Additionally, the presence of the negative strand of the clone was consistently detected in the IE1-SBV-transfected Sf9 cells and injected pupae of both
A. mellifera and
A. cerana, suggesting the active replication of the cloned virus in these hosts. Although the possibility of false-positive results in the negative strand detection cannot be completely ruled out [
32], our repeated transfections of Sf9 cells consistently yielded the same findings, indicating active replication within the cells. Moreover, the use of DNA plasmids with the IE1 promoter streamlines the sequence editing processes of bee-infecting RNA viruses and offers Sf9 cells and similar cell lines as valuable tools for evaluating the viability of edited clones, reducing the need for repetitive sequence editing and bioassays using honey bees. The synthesis of bee-infecting RNA virus clones using the IE1 promoter holds potential for facilitating reverse genetic studies, thereby enhancing our understanding of bee-infecting RNA virus biology and pathology.
A few limitations of using Sf9 cells, not the natural host, as a surrogate host for SBV have been revealed. Previous studies [
21,
33], which used purified virions and viral genome RNA, respectively, suggested the potential of SBV to infect surrogate cell lines. However, these studies reported failures to inoculate naïve cells upon subculture and similar findings were observed in a study on Deformed wing virus in Sf9 cells [
34]. In this study, we also noted that the cells with the expressing tag of the SBV clone are not increased with a prolonged incubation time. Since the expressing tag is stable within the SBV clone [
14], this observation suggested that the clone cannot disseminate to adjacent cells naturally or form infective virions. This phenomenon also explained the low count in the RT-qPCR for SBV genome copies of the transfected Sf9 cells, because only the cells transfected with the plasmid synthesized the SBV viral RNA. In contrast, an Iflavirus found in lepidopteran,
Perina nuda virus, PnV [
35], achieved a nearly 100% infection rate in Sf9 cells and produced millions of viruses. While Sf9 cells may not be ideal hosts for massive cloned virus production in bee-infecting RNA viruses, they can still be valuable for assessing whether the clone is capable of replicating, which is a crucial step in trials involving honey bees. By utilizing this approach, the need for extensive resources when testing multiple mutated clones simultaneously can be significantly reduced.
The confocal microscope observations of the transfected Sf9 cells illustrated obvious alterations in membrane structures within the cytoplasm. This result is consistent with a lipidomic study using
A. cerana larvae [
36], which found SBV infection significantly altered the host lipid profiles. Since the synthesized SBV clones did not disseminate to naïve Sf9 cells, these membrane structure alterations were probably not caused by the entry of SBV, which may involve an endocytosis [
37]. Additional TEM observation (
Figure S2) suggested these membrane structures are mitochondria, ER-like vesicles, and multi-membrane vesicles. However, based on the current results, we cannot determine if the membrane structures are involved in the antiviral responses of the cell [
38,
39] or intermediate vesicles forming extracellular vesicles [
40,
41].
The transfection method for the plasmid containing the viral full-length cDNA genome has not been fully optimized in this study. The transfection rate of the plasmids in Sf9 cells was found to be lower than 10%, which did not meet our initial expectations. However, this estimation might be an overestimate, as we only counted areas containing cells with fluorescence. Prior experiences with Lipofectamine-mediated transfections of bacmids containing a baculovirus genome of over 100 kb in size into Sf9 cells were successful; however, those experiments involved enrichment through subcultures, a process not feasible for the infectious clone in this study. Furthermore, the suggested volumes on the Lipofectamine manufacturer’s protocol were optimized for cell cultures using large volumes—specifically, 500 microliters with 4000 ng plasmids for half a million seeded cells. This suggested volume is unsuitable for honey bee pupa injection, as a honey bee pupa consists of far more cells but can only accommodate approximately 10 microliters of injection before the body cavity becomes severely distended. Consequently, we had to use much higher concentrated plasmids (1800 and 2500 ng in eight microliters final mixture) mixed with Lipofectamine. The high concentrations of plasmid DNA and liposomes may have affected their interaction, and we found that injecting 5000 ng of the plasmid did not yield more infection when the Lipofectamine was not increased. However, our application and optimization of the transfection and injection method were limited by our lack of expertise in such protocols. Exploring other transfection reagents or methods that are suitable for delivering large DNA fragments within relatively small volumes may enhance the results in cells and honey bees. Additionally, the choice of the expressing tag, EGFP, may not be the most suitable option for honey bee pupae due to the confusion with autofluorescence from the control pupae. Other alternatives, such as mCherry or nanoluc [
42], may prove more effective for pupa studies.
The observed infection rate differences in pupae may be attributed to their anti-virus immune responses. The consistently low infection rates in Sf9 cells during transfection suggest that only a small number of cells within the injected pupae were successfully transfected by the plasmid, leading to the synthesis of infectious clones in limited quantities that may not have been sufficient to overcome the host’s anti-virus defenses. The results of plasmid injections in both
A. mellifera and
A. cerana pupae support the hypothesis that host immune responses may restrict the infection success of the injected clones.
A. cerana pupae consistently displayed a 75% infection rate with 2500 ng plasmid in repetition trials using pupae from different colonies, while
A. mellifera showed only a 25% infection rate with the same dosage. Similarly, another previous injection of
A. mellifera using 1800 ng plasmid yielded a comparable infection rate (33%) in trials. A direct comparison between
A. cerana and
A. mellifera injections indicates that
A. cerana is approximately three times more susceptible to the SBV clone. This finding gains further significance from the fact that
A. cerana colonies exhibited a higher susceptibility to SBV in the field [
43]. Interestingly, the observation of highly infected individual pupae was specific to the
A. cerana pupae, suggesting some individuals in the colony were highly vulnerable to the virus. Notably, these
A. cerana colonies had previous pre-exposures to the SBV variant during our continuous survey, which may have resulted in primed immune responses to the SBV variant and could explain the high deviations of the infection intensities; however, their susceptibility remained much higher than that of
A. mellifera. The identified susceptibility difference between
A. cerana and
A. mellifera in this study necessitates further investigation with larger sample sizes, well-defined genetic backgrounds, and pre-exposures to the virus. Additional research is needed to better understand the underlying factors contributing to the differential susceptibility of
A. cerana and
A. mellifera to the SBV variant.
Our successful generation of infectious clones in Sf9 cells and honey bee pupae offers new possibilities for testing the viability and virulence of edited clones, reducing the time and labor involved in these critical investigations. Our approach presents an alternative in situations where bees free of the cloned virus species are unavailable in the field, as cultured cells can serve as surrogate hosts. As we consider future applications, the use of honey bee cell lines [
44] or primary cultures [
45] may be explored since the same plasmid can drive the clone synthesis in honey bee pupae. However, caution should be exercised regarding potential virus contamination concerns in these cultures. Continued research into the membrane structures observed in Sf9 cell cytoplasm and an exploration of alternative expressing tags for pupae studies will undoubtedly advance our understanding of bee-infecting RNA virus dynamics.
In conclusion, our study has successfully demonstrated the synthesis of SBV clones using the IE1 promoter in both Sf9 cells and honey bee pupae. Despite some limitations in the transfection method and the challenges posed by hosts, our findings hold promise for streamlining reverse genetic studies of bee-infecting RNA viruses. By utilizing DNA plasmids with the IE1 promoter, we have achieved the efficient expression and replication of the cloned virus in laboratory-controlled settings. These advancements offer a valuable tool for investigating the biology and pathology of bee-infecting RNA viruses in a more efficient and controlled manner. Overall, this study offers significant potential to advance our knowledge of bee–virus interactions and may ultimately contribute to the development of effective strategies for safeguarding the health of these critical pollinators. We hope that our work will inspire further research in this area and contribute to the collective efforts aimed at securing a healthy future for these essential pollinators.











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}