
Touching Base with Some Mediterranean Diseases of Interest from Paradigmatic Cases at the “Magna Graecia” University Unit of Infectious Diseases: A Didascalic Review

Abstract
:1. Introduction
1.1. Definition of Zoonoses and Main Components
1.2. Transmission Mechanisms and Selected Zoonoses
- Rickettsiosis;
- Ehrlichiosis;
- Borreliosis;
- Bartonellosis;
- Brucellosis;
- Leishmaniasis.
2. Zoonoses with Epidemiological Relevance in the Mediterranean Area
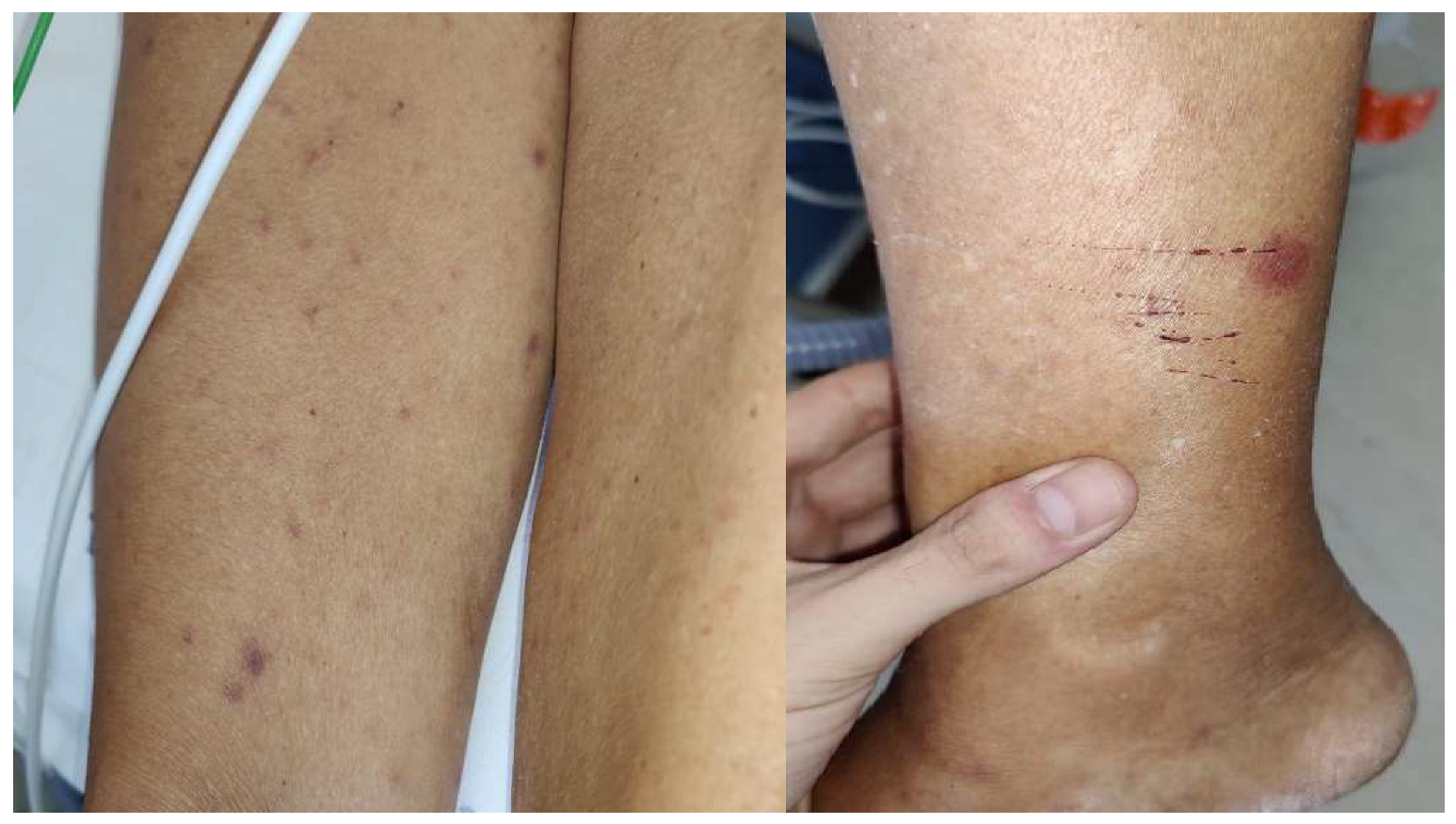
2.1. Rickettsiosis
- Spotted fever (to which R. rickettsii, R. conorii, R. parkeri, and several others belong);
- Typhus (R. prowazekii and R. typhi);
- Ancestral (R. bellii and R. canadensis, even if not known to be pathogenic);
- Transitional (R. akari, R. australis, and R. felis) [14].
- Recruitment of C6–9 proteins to produce a lytic pore to kill the microorganism;
- Opsonization from the deposition of C3b and C5b to improve phagocytic cell digestion;
- Release of anaphylatoxins C3a and C5a, which recruit immune cells to areas of complement activation [31].
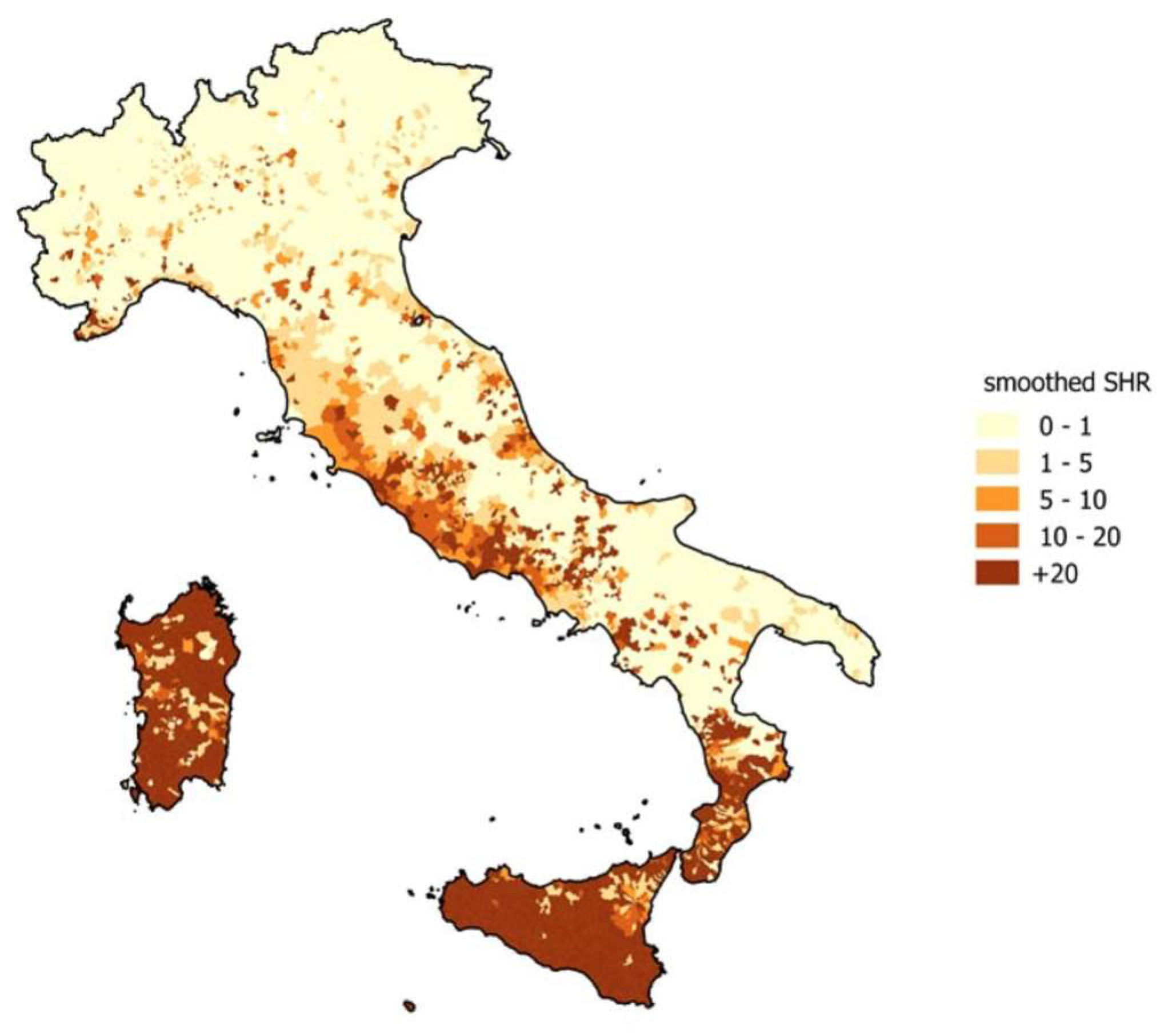
2.2. Brucellosis
- B. melitensis (isolated from ruminants such as goats, sheep, or camels);
- B. abortus (isolated from livestock);
- B. suis (isolated from pigs);
- Those with objective evidence of a Brucella infection (bacterium isolated from tissue, blood culture, or specific antibody titers) and/or focal complications (such as osteomyelitis, spondylitis, or uveitis) [39];
- Those with persistent symptoms in the absence of objective infection evidence (serological values or positive cultures); in this case, the symptoms may include general malaise, psychiatric disorders (depression, anxiety, and emotional lability), tremor or arthralgia, insomnia, or sexual disturbances [39].
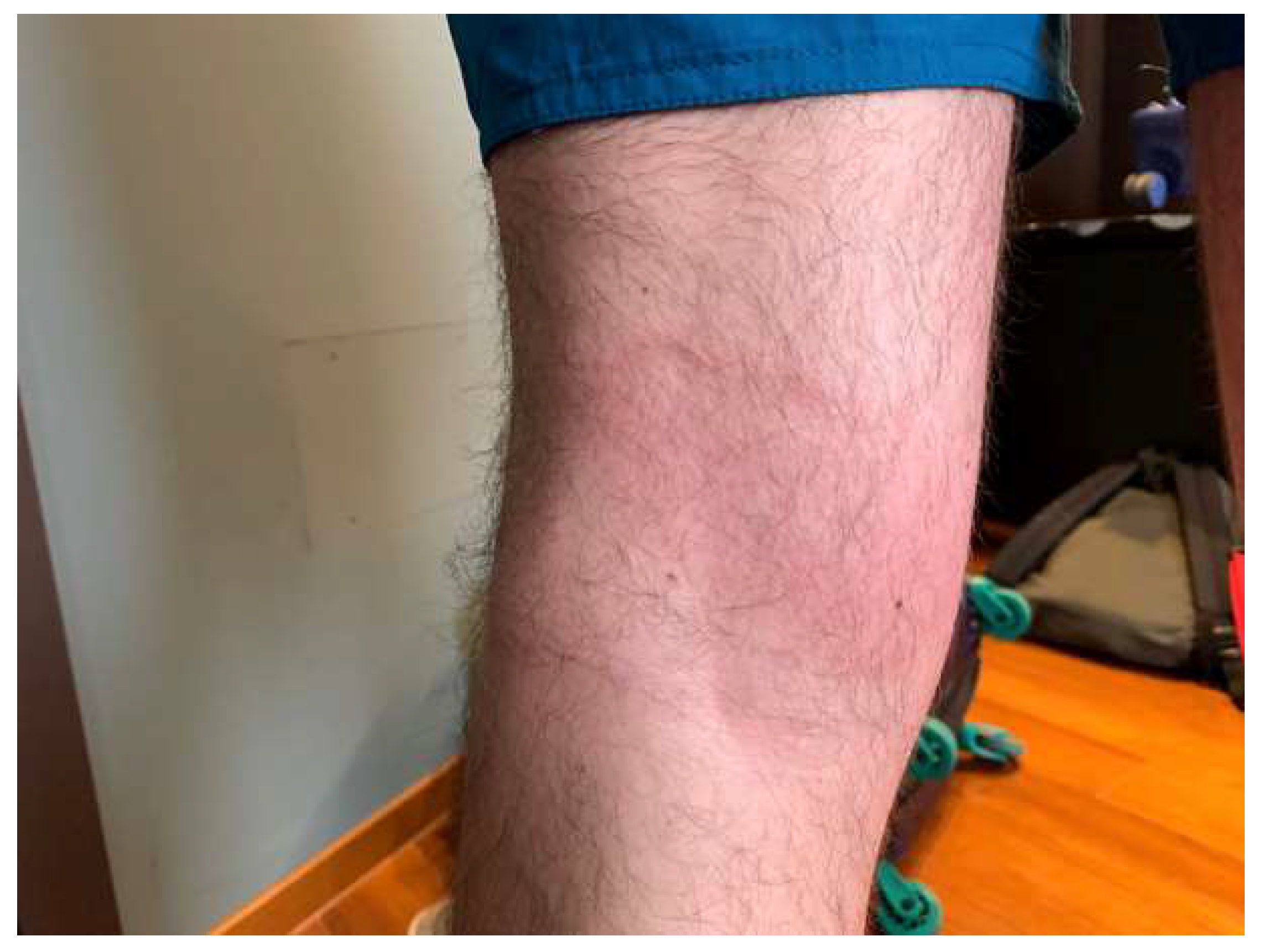
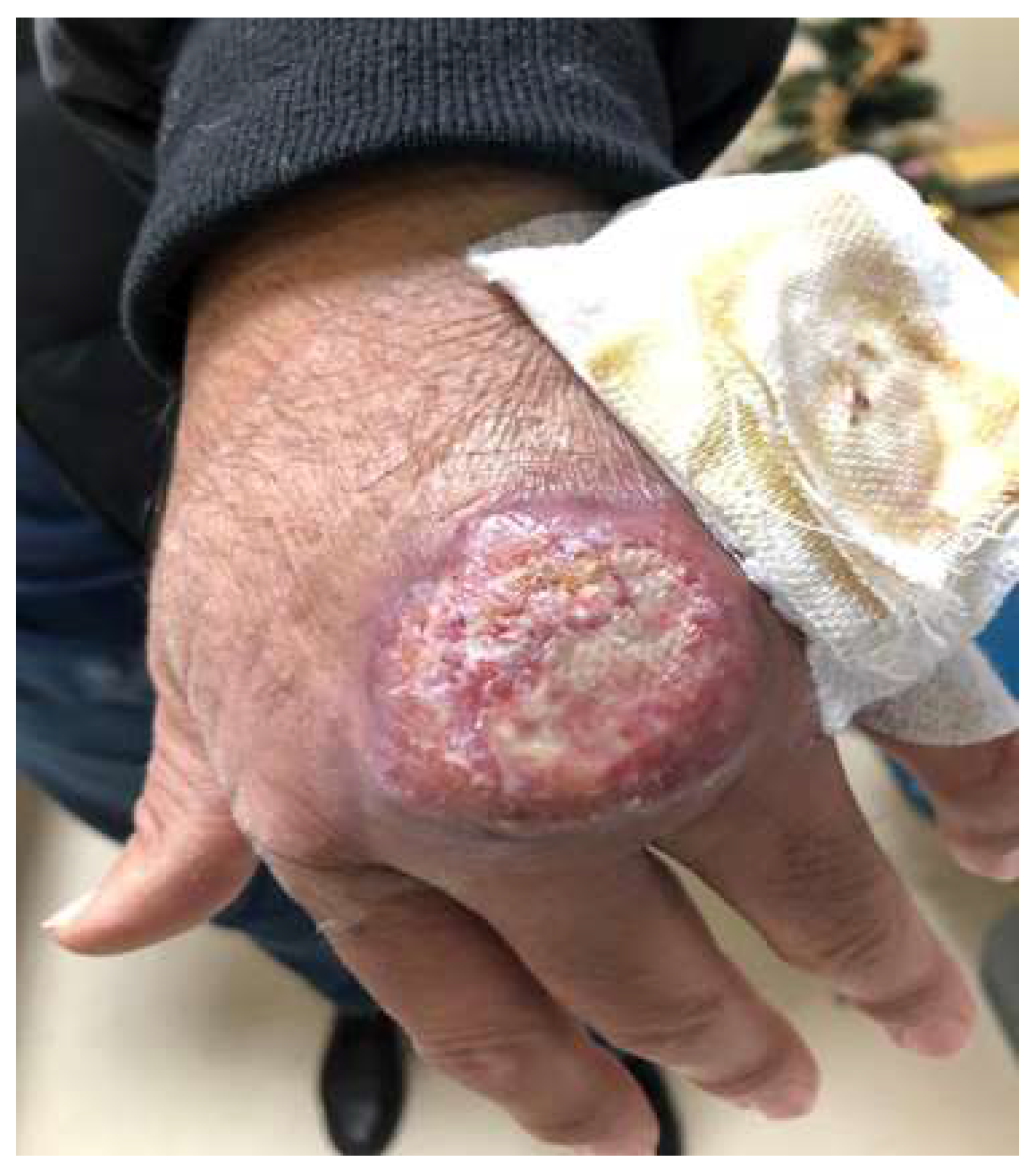
2.3. Bartonellosis
2.4. Ehrlichiosis
- Neoehrlichia: Candidatus N. mikurensis and N. lotoris;
- Ehrlichia: E. minasensis, E. ruminantium, E. canis, E. muris subsp muris, E. muris subsp eauclairensis, E. chaffeensis, the Panola Mountain Ehrlichia, and E. ewingii;
- Anaplasma: A. phagocytophilum, A. marginale, A. centrale, A. ovis, A. platys, A. bovis, A. capra, and Aegyptianella pullorum [82].
2.5. Lyme Borreliosis
- Joint involvement with swelling and pain, particularly in the large joints; the knee joint is affected with a pattern of oligoarticular arthritis that can evolve to severe chronic arthritic lesions [111].
- Cardiac involvement with acute onset of intermittent atrioventricular heart block, which is sometimes associated with clinical evidence of myopericarditis [112].
- Neurological involvement including meningitis, unilateral facial palsy, other cranial neuritis, and radiculitis may occur. Rare manifestations of the central nervous system include encephalomyelitis and chronic meningitis [111].
- Borrelial lymphocytoma is a rare skin manifestation (mainly seen in Europe) that presents as a solitary swelling most frequently located on the ear lobe and in the region of the areola mammae [112].
- Acrodermatitis chronica atrophicans is a skin manifestation exclusively observed in Europe, often seen on acral parts of the body and on the extensor part of hands or feet, and initially is usually unilateral; later, it may become more or less symmetrical with thin and wrinkled skin [112].
2.6. Leishmaniasis
3. Future Perspectives and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alho, A.M.; Lima, C.; Colella, V.; de Carvalho, L.M.; Otranto, D.; Cardoso, L. Awareness of zoonotic diseases and parasite control practices: A survey of dog and cat owners in Qatar. Parasites Vectors 2018, 11, 1–7. [Google Scholar] [CrossRef]
- Jánová, E. Emerging and threatening vector-borne zoonoses in the world and in Europe: A brief update. Pathog. Glob Health. 2019, 113, 49–57. [Google Scholar] [PubMed]
- McArthur, D.B. Emerging infectious diseases. Nurs. Clin. 2019, 54, 297–311. [Google Scholar]
- Available online: https://www.epicentro.iss.it/zecche/ (accessed on 17 July 2022).
- Samuel, G.H.; Adelman, Z.N.; Myles, K.M. Temperature-dependent effects on the replication and transmission of arthropod-borne viruses in their insect hosts. Curr. Opin. Insect Sci. 2016, 16, 108–113. [Google Scholar] [CrossRef]
- Eisen, R.J.; Kugeler, K.J.; Eisen, L.; Beard, C.B.; Paddock, C.D. Tick-Borne Zoonoses in the United States: Persistent and Emerging Threats to Human Health. ILAR J. 2017, 58, 319–335. [Google Scholar] [CrossRef] [PubMed]
- Horka, H.; Staudt, V.; Klein, M.; Taube, C.; Reuter, S.; Dehzad, N.; Schmitt, E. The tick salivary protein sialostatin L inhibits the Th9-derived production of the asthma-promoting cytokine IL-9 and is effective in the prevention of experimental asthma. J. Immunol. 2012, 188, 2669–2676. [Google Scholar] [PubMed]
- Sultana, H.; Neelakanta, G. Arthropod exosomes as bubbles with message(s) to transmit vector-borne diseases. Curr. Opin. Insect Sci. 2020, 40, 39–47. [Google Scholar]
- Neelakanta, G.; Sultana, H. Tick saliva and salivary glands: What do we know so far on their role in arthropod blood feeding and pathogen transmission. Front. Cell. Infect. Microbiol. 2022, 11, 816547. [Google Scholar] [PubMed]
- Morelli, A.; Duranti, A.; Pauri, P.; Secondini, S. Laboratory: A Privileged Point of View in Zoonoses Surveil-Lance. 2016. Available online: http://spvet.it/indice-spv.html#Numero99 (accessed on 20 July 2022).
- Sgroi, G.; Iatta, R.; Lovreglio, P.; Stufano, A.; Laidoudi, Y.; Mendoza-Roldan, J.A.; Otranto, D. Detection of Endosymbiont Candidatus Midichloria mitochondrii and tickborne pathogens in humans exposed to tick bites, Italy. Emerg. Infect. Dis. 2022, 28, 1824. [Google Scholar]
- Osterloh, A. Immune response against rickettsiae: Lessons from murine infection models. Med. Microbiol. Immunol. 2017, 206, 403–417. [Google Scholar]
- Milagres, B.S.; Gomes, G.G.; Galvao, M.A.M.; Freitas, R.N.; Pacheco, R.; Bouyer, D.H.; Montandon, C.E.; Silveira, I.; Barcelos, R.M.; Mafra, C.L.; et al. Rickettsia in Synanthropic and Domestic Animals and Their Hosts from Two Areas of Low Endemicity for Brazilian Spotted Fever in the Eastern Region of Minas Gerais, Brazil. Am. J. Trop. Med. Hyg. 2010, 83, 1305–1307. [Google Scholar] [CrossRef]
- Available online: https://www.epicentro.iss.it/zecche/FebbreBottonosa2001-2015 (accessed on 17 July 2022).
- Available online: https://www.epicentro.iss.it/zecche/rickettsiosi (accessed on 21 July 2022).
- Heyman, P.; Cochez, C.; Hofhuis, A.; Van Der Giessen, J.; Sprong, H.; Porter, S.R.; Papa, A. A clear and present danger: Tick-borne diseases in Europe. Exp. Rev. Anti-Infect. Ther. 2010, 8, 33–50. [Google Scholar] [CrossRef]
- Creus, B.F.; Cueto, F.B.; Arenas, E.E.; Sanahuja, R.V.; Espin, T.M.; Salas, M.N.; Borrell, A.C.; Cuesta, J.M.; Porta, F.S. Mediterranean Spotted Fever: A Cooperative Study of 227 Cases. Clin. Infect. Dis. 1985, 7, 635–642. [Google Scholar] [CrossRef]
- Mert, A.; Ozaras, R.; Tabak, F.; Bilir, M.; Ozturk, R. Mediterranean spotted fever: A review of fifteen cases. J. Dermatol. 2006, 33, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Rovery, C.; Brouqui, P.; Raoult, D. Questions on Mediterranean Spotted Fever a Century after Its Discovery. Emerg. Infect. Dis. 2008, 14, 1360–1367. [Google Scholar] [CrossRef] [PubMed]
- Parola, P.; Paddock, C.D.; Socolovschi, C.; Labruna, M.B.; Mediannikov, O.; Kernif, T.; Raoult, D. Update on tick-borne rickettsioses around the world: A geographic approach. Clin. Microbiol. Rev. 2013, 26, 657–702. [Google Scholar] [CrossRef]
- Faccini-Martínez, A.; García-Álvarez, L.; Hidalgo, M.; Oteo, J.A. Syndromic classification of rickettsioses: An approach for clinical practice. Int. J. Infect. Dis. 2014, 28, 126–139. [Google Scholar] [CrossRef]
- Chapman, A.S. Diagnosis and management of tickborne rickettsial diseases: Rocky Mountain spotted fever, ehrlichioses, and anaplasmo-sis-United States: A practical guide for physicians and other health-care and public health professionals. MMWR Recomm. Rep. 2006, 65, 1–44. [Google Scholar]
- McQuiston, J.H.; Dunn, J.; Morris, K.; Carpenter, L.R.; Moncayo, A.C.; Chung, I.; McElroy, K.; Wiedeman, C.; Porter, S.; Kato, C.; et al. Inadequacy of IgM Antibody Tests for Diagnosis of Rocky Mountain Spotted Fever. Am. J. Trop. Med. Hyg. 2014, 91, 767–770. [Google Scholar] [CrossRef] [PubMed]
- La Scola, B.; Raoult, D. Laboratory diagnosis of rickettsioses: Current approaches to diagnosis of old and new rickettsial diseases. J. Clin. Microbiol. 1997, 35, 2715–2727. [Google Scholar] [CrossRef]
- Paris, D.H.; Dumler, J.S. State of the art of diagnosis of rickettsial diseases: The use of blood specimens for diagnosis of scrub typhus, spotted fever group rickettsiosis, and murine typhus. Curr. Opin. Infect. Dis. 2016, 29, 433. [Google Scholar] [CrossRef]
- Raoult, D.; Dupont, H.T.; Caraco, P.; Brouqui, P.; Drancourt, M.; Charrel, C. Mediterranean spotted fever in Marseille: Descriptive epidemiology and the influence of climatic factors. Eur. J. Epidemiol. 1992, 8, 192–197. [Google Scholar] [CrossRef] [PubMed]
- Silva-Ramos, C.R.; Hidalgo, M.; Faccini-Martínez, A. Clinical, epidemiological, and laboratory features of Rickettsia parkeri rickettsiosis: A systematic review. Ticks Tick-Borne Dis. 2021, 12, 101734. [Google Scholar] [CrossRef]
- Helminiak, L.; Mishra, S.; Kim, H.K. Pathogenicity and virulence of Rickettsia. Virulence 2022, 13, 1752–1771. [Google Scholar] [CrossRef] [PubMed]
- Papp, S.; Moderzynski, K.; Rauch, J.; Heine, L.; Kuehl, S.; Richardt, U.; Mueller, H.; Fleischer, B.; Osterloh, A. Liver Necrosis and Lethal Systemic Inflammation in a Murine Model of Rickettsia typhi Infection: Role of Neutrophils, Macrophages and NK Cells. PLOS Neglected Trop. Dis. 2016, 10, e0004935. [Google Scholar] [CrossRef] [PubMed]
- Janeway, C.A., Jr.; Travers, P.; Walport, M.; Shlomchik, M.J. Immunobiology: The Immune System in Health and Disease. 2001. Available online: https://www.ncbi.nlm.nih.gov/books/NBK27100/ (accessed on 17 July 2022).
- Ricklin, D.; Reis, E.S.; Lambris, J.D. Complement in disease: A defence system turning offensive. Nat. Rev. Nephrol. 2016, 12, 383–401. [Google Scholar] [CrossRef]
- Dahmani, M.; Cook, J.H.; Zhu, J.C.; Riley, S.P. Contribution of classical complement activation and IgM to the control of Rickettsia infection. Mol. Microbiol. 2021, 116, 1476–1488. [Google Scholar] [CrossRef]
- Klein, M.; Brühl, T.-J.; Staudt, V.; Reuter, S.; Grebe, N.; Gerlitzki, B.; Hoffmann, M.; Bohn, T.; Ulges, A.; Stergiou, N.; et al. Tick Salivary Sialostatin L Represses the Initiation of Immune Responses by Targeting IRF4-Dependent Transcription in Murine Mast Cells. J. Immunol. 2015, 195, 621–631. [Google Scholar] [CrossRef]
- Koch, S.; Sopel, N.; Finotto, S. Th9 and other IL-9-producing cells in allergic asthma. Semin. Immunopathol. 2017, 39, 55–68. [Google Scholar] [CrossRef]
- Tete, S.; Saggini, A.; Maccauro, G.; Rosati, M.; Conti, F.; Cianchetti, E.; Tripodi, D.; Toniato, E.; Fulcheri, M.; Salini, V.; et al. Interleukin-9 and mast cells. J. Biol. Regul. Homeost. Agents 2012, 26, 432–445. [Google Scholar]
- Available online: https://www.epicentro.iss.it/brucellosi/epidemiologia-italia (accessed on 23 July 2022).
- Godfroid, J.; Cloeckaert, A.; Liautard, J.P.; Kohler, S.; Fretin, D.; Walravens, K.; Letesson, J.J. From the discovery of the Malta fever’s agent to the discovery of a marine mammal reservoir, brucellosis has continuously been a re-emerging zoonosis. Vet. Res. 2005, 36, 313–326. [Google Scholar] [CrossRef]
- Seleem, M.; Boyle, S.M.; Sriranganathan, N. Brucellosis: A re-emerging zoonosis. Veter. Microbiol. 2010, 140, 392–398. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.uptodate.com/contents/brucellosis-epidemiology-microbiology-clinical-manifestations-and-diagnosis (accessed on 30 July 2022).
- Available online: https://www.ecdc.europa.eu/en/brucellosis (accessed on 30 July 2022).
- Pappas, G.; Aritidis, N.; Bosilovsi, M.; Tsianos, E. Brucellosis. N. Engl. J. Med. 2005, 352, 2325–2336. [Google Scholar] [CrossRef] [PubMed]
- Harrison, E.R.; Posada, R. Brucellosis. Pediatr. Rev. 2018, 39, 222–224. [Google Scholar] [CrossRef]
- Bukhari, E. Pediatric brucellosis: An update review for the new millennium. Saudi Med. J. 2018, 39, 336–341. [Google Scholar] [CrossRef]
- Franco, M.P.; Mulder, M.; Gilman, R.H.; Smits, H.L. Human brucellosis. Lancet Infect. Dis. 2007, 7, 775–786. [Google Scholar] [CrossRef]
- Solera, J.; Martinez-Alfaro, E.; Espinosa, A. Recognition and Optimum Treatment of Brucellosis. Drugs 1997, 53, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Brucellosis Reference Guide Centers for Disease Control (CDC). Available online: https://www.cdc.gov/brucellosis/pdf/brucellosi-reference-guide.pdf (accessed on 14 August 2023).
- Al Dahouk, S.; Tomaso, H.; Nöckler, K.; Neubauer, H.; Frangoulidis, D. Laboratory-based diagnosis of brucellosis—A review of the literature. Part II: Serological tests for brucellosis. Clin. Lab. 2003, 49, 577–589. [Google Scholar]
- de los Ángeles Mantecón, M.; Gutiérrez, M.P.; del Pilar Zarzosa, M.; Fernández-Lago, L.; de Dios Colmenero, J.; Vizcaíno, N.; Bratos, M.A.; Almaraz, A.; Cubero, A.; Muñoz, M.F.; et al. Influence of brucellosis history on serological diagnosis and evolution of patients with acute brucellosis. J. Infect. 2008, 57, 397–403. [Google Scholar] [CrossRef]
- Yagupsky, P.; Morata, P.; Colmenero, J.D. Laboratory Diagnosis of Human Brucellosis. Clin. Microbiol. Rev. 2019, 33, 1. [Google Scholar] [CrossRef]
- López-Santiago, R.; Sánchez-Argáez, A.B.; De Alba-Núñez, L.G.; Baltierra-Uribe, S.L.; Moreno-Lafont, M.C. Immune Response to Mucosal Brucella Infection. Front. Immunol. 2019, 10, 1759. [Google Scholar] [CrossRef]
- Muñoz González, F.; Sycz, G.; Alonso Paiva, I.M.; Linke, D.; Zorreguieta, A.; Baldi, P.C.; Ferrero, M.C. The BtaF adhesin is necessary for full virulence during respiratory infection by Brucella suis and is a novel immunogen for nasal vaccination against Brucella infection. Front. Immunol. 2019, 10, 1775. [Google Scholar] [CrossRef]
- Bialer, M.G.; Sycz, G.; González, F.M.; Ferrero, M.C.; Baldi, P.C.; Zorreguieta, A. Adhesins of Brucella: Their Roles in the Interaction with the Host. Pathogens 2020, 9, 942. [Google Scholar] [CrossRef]
- Ferrero, M.C.; Fossati, C.A.; Rumbo, M.; Baldi, P.C. Brucella invasion of human intestinal epithelial cells elicits a weak proinflammatory response but a significant CCL20 secretion. FEMS Immunol. Med. Microbiol. 2012, 66, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Paixao, T.A.; Roux, C.M.; den Hartigh, A.B.; Sankaran-Walters, S.; Dandekar, S.; Santos, R.L.; Tsolis, R.M. Establishment of systemic Brucella melitensis infection through the digestive tract requires urease, the type IV secretion system, and lipopolysaccharide O antigen. Infect. Immun. 2009, 77, 4197–4208. [Google Scholar] [CrossRef] [PubMed]
- Copin, R.; Vitry, M.-A.; Mambres, D.H.; Machelart, A.; De Trez, C.; Vanderwinden, J.-M.; Magez, S.; Akira, S.; Ryffel, B.; Carlier, Y.; et al. In Situ Microscopy Analysis Reveals Local Innate Immune Response Developed around Brucella Infected Cells in Resistant and Susceptible Mice. PLOS Pathog. 2012, 8, e1002575. [Google Scholar] [CrossRef]
- Vitry, M.A.; Hanot Mambres, D.; De Trez, C.; Akira, S.; Ryffel, B.; Letesson, J.J.; Muraille, E. Humoral immunity and CD4+ Th1 cells are both necessary for a fully protective immune response upon secondary infection with Brucella melitensis. J. Immunol. 2014, 192, 3740–3752. [Google Scholar] [CrossRef]
- Ives, A.; Masina, S.; Castiglioni, P.; Prevel, F.; Revaz-Breton, M.; Hartley, M.A.; Ronet, C. MyD88 and TLR9 dependent immune responses mediate resistance to Leishmania guyanensis infections, irrespective of Leishmania RNA virus burden. PLoS ONE 2014, 9, e96766. [Google Scholar] [CrossRef]
- Deqiu, S.; Donglou, X.; Jiming, Y. Epidemiology and control of brucellosis in China. Veter Microbiol. 2002, 90, 165–182. [Google Scholar] [CrossRef] [PubMed]
- Abkar, M.; Fasihi-Ramandi, M.; Kooshki, H.; Lotfi, A.S. Intraperitoneal immunization with Urease loaded N-trimethyl Chitosan nanoparticles elicits high protection against Brucella melitensis and Brucella abortus infections. Immunol. Lett. 2018, 199, 53–60. [Google Scholar] [CrossRef]
- Mazzitelli, M.; Lamberti, A.G.; Quirino, A.; Marascio, N.; Barreca, G.S.; Costa, C.; Torti, C. Utility of molecular identification and quantitation of Bartonella species with species-specific real-time PCR for monitoring treatment response: A case series. Open Microbiol. J. 2018, 12, 148. [Google Scholar] [CrossRef]
- Mosbacher, M.E.; Klotz, S.; Klotz, J.; Pinnas, J.L. Bartonella henselae and the potential for arthropod vector-borne transmission. Vector-Borne Zoonot. Dis. 2011, 11, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Chomel, B.B.; Boulouis, H.-J.; Maruyama, S.; Breitschwerdt, E.B. Bartonella Spp. in Pets and Effect on Human Health. Emerg. Infect. Dis. 2006, 12, 389–394. [Google Scholar] [CrossRef]
- Graziani, C.; Duranti, A.; Morelli, A.; Busani, L.; Pezzotti, P. Zoonosi in Italia nel Periodo 2009–2013. Roma: Istituto Superiore di Sanità. 2016. 2016. Rapporti ISTISAN 16/1. Available online: https://www.iss.it/rapporti-istisan/-/asset_publisher/Ga8fOpve0fNN/content/id/5183699 (accessed on 17 July 2022).
- Bos, F.; Chauveau, B.; Ruel, J.; Fontant, G.; Campistron, E.; Meunier, C.; Jambon, F.; Moreau, K.; Delmas, Y.; Couzi, L.; et al. Serious and Atypical Presentations of Bartonella henselae Infection in Kidney Transplant Recipients. Open Forum Infect. Dis. 2022, 9, ofac059. [Google Scholar] [CrossRef] [PubMed]
- Luciani, L.; El Baroudi, Y.; Prudent, E.; Raoult, D.; Fournier, P.-E. Bartonella infections diagnosed in the French reference center, 2014–2019, and focus on infections in the immunocompromised. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 2407–2410. [Google Scholar] [CrossRef]
- Mosepele, M.; Mazo, D.; Cohn, J. Bartonella Infection in Immunocompromised Hosts: Immunology of Vascular Infection and Vasoproliferation. J. Immunol. Res. 2011, 2012, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Karem, K.L.; Paddock, C.D.; Regnery, R.L. Bartonella henselae, B. quintana, and B. bacilliformis: Historical pathogens of emerging significance. Microbes Infect. 2000, 2, 1193–1205. [Google Scholar] [CrossRef] [PubMed]
- Raoult, D.; Roux, V. The Body Louse as a Vector of Reemerging Human Diseases. Clin. Infect. Dis. 1999, 29, 888–911. [Google Scholar] [CrossRef]
- Foucault, C.; Brouqui, P.; Raoult, D. Bartonella quintana Characteristics and Clinical Management. Emerg Infect Dis. 2006, 12, 217–223. [Google Scholar] [CrossRef]
- Maurin, M.; Raoult, D. Bartonella (Rochalimaea) quintana infections. Clin. Microbiol. Rev. 1996, 9, 273–292. [Google Scholar] [CrossRef]
- Available online: https://www.uptodate.com/contents/clinical-features-diagnosis-and-treatment-of-bartonella-quintana-infections (accessed on 3 August 2022).
- Ohl, M.E.; Spach, D.H. Bartonella quintana and Urban Trench Fever. Clin. Infect. Dis. 2000, 31, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Spach, D.H.; Koehler, J.E. Bartonella-associated infections. Infect. Dis. Clin. 1998, 12, 137–155. [Google Scholar] [CrossRef] [PubMed]
- Jacomo, V.; Kelly, P.J.; Raoult, D. Natural History of Bartonella Infections (an Exception to Koch’s Postulate). Clin. Vaccine Immunol. 2002, 9, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Carithers, H.A. Cat-scratch disease: An overview based on a study of 1200 patients. Am. J. Dis. Child. 1985, 139, 1124–1133. [Google Scholar] [CrossRef]
- Charles, R.C.; Sertic, M.; Neilan, A.M.; Sohani, A.R. Case 11-2021: A 39-year-old woman with fever, flank pain, and inguinal lymphadenopathy. N. Engl. J. Med. 2021, 384, 1448–1456. [Google Scholar] [CrossRef]
- Paddock, C.D.; Childs, J.E. Ehrlichia chaffeensis: A Prototypical Emerging Pathogen. Clin. Microbiol. Rev. 2003, 16, 37–64. [Google Scholar] [CrossRef]
- Angelakis, E.; Raoult, D. Pathogenicity and treatment of Bartonella infections. Int. J. Antimicrob. Agents 2014, 44, 16–25. [Google Scholar] [CrossRef]
- Choi, E.W.; Lee, H.W.; Lee, J.S.; Kim, I.Y.; Shin, J.H.; Seong, J.K. Ahnak-knockout mice show susceptibility to Bartonella henselae infection because of CD4+ T cell inactivation and decreased cytokine secretion. BMB Rep. 2019, 52, 289. [Google Scholar] [CrossRef]
- Harms, A.; Dehio, C. Intruders below the Radar: Molecular Pathogenesis of Bartonella spp. Clin. Microbiol. Rev. 2012, 25, 42–78. [Google Scholar] [CrossRef]
- Matza, D.; Badou, A.; Kobayashi, K.S.; Goldsmith-Pestana, K.; Masuda, Y.; Komuro, A.; McMahon-Pratt, D.; Marchesi, V.T.; Flavell, R.A. A Scaffold Protein, AHNAK1, Is Required for Calcium Signaling during T Cell Activation. Immunity 2008, 28, 64–74. [Google Scholar] [CrossRef]
- Available online: https://www.uptodate.com/contents/human-ehrlichiosis-and-anaplasmosis (accessed on 4 August 2022).
- Dumler, J.S.; Choi, K.-S.; Garcia-Garcia, J.C.; Barat, N.S.; Scorpio, D.G.; Garyu, J.W.; Grab, D.J.; Bakken, J.S. Human Granulocytic Anaplasmosis and Anaplasma phagocytophilum. Emerg. Infect. Dis. 2005, 11, 1828–1834. [Google Scholar] [CrossRef] [PubMed]
- Dumler, J.S.; Madigan, J.E.; Pusterla, N.; Bakken, J.S. Ehrlichioses in Humans: Epidemiology, Clinical Presentation, Diagnosis, and Treatment. Clin. Infect. Dis. 2007, 45, S45–S51. [Google Scholar] [CrossRef] [PubMed]
- Guccione, C.; Colomba, C.; Tolomeo, M.; Trizzino, M.; Iaria, C.; Cascio, A. Rickettsiales in Italy. Pathogens 2021, 10, 181. [Google Scholar] [CrossRef] [PubMed]
- Heitman, K.N.; Dahlgren, F.S.; Drexler, N.A.; Massung, R.F.; Behravesh, C.B. Increasing incidence of ehrlichiosis in the United States: A summary of national surveillance of Ehrlichia chaffeensis and Ehrlichia ewingii infections in the United States, 2008–2012. Am. J. Trop. Med. Hyg. 2016, 94, 52. [Google Scholar] [CrossRef] [PubMed]
- Moniuszko-Malinowska, A.; Dunaj, J.; Andersson, M.O.; Czupryna, P.; Zajkowska, J.; Guziejko, K.; Pancewicz, S. Assessment of Anaplasma phagocytophilum presence in early Lyme borreliosis manifested by erythema migrans skin lesions. Travel Med. Infect. Dis. 2020, 36, 101648. [Google Scholar] [CrossRef]
- Moniuszko-Malinowska, A.; Dunaj, J.; Andersson, M.O.; Chmielewski, T.; Czupryna, P.; Groth, M.; Pancewicz, S. Anaplasmosis in Poland–analysis of 120 patients. Ticks Tick-borne Dis. 2021, 12, 101763. [Google Scholar] [CrossRef]
- Ehrlichiosis and Anaplasmosis 2008 Case Definition. United States Centers for Disease Control. Available online: https://ndc.services.cdc.gov/case-definitions/ehrlichiosis-and-anaplasmosis-2008/ (accessed on 9 August 2023).
- Blanco, J.; Oteo, J. Human granulocytic ehrlichiosis in Europe. Clin. Microbiol. Infect. 2002, 8, 763–772. [Google Scholar] [CrossRef]
- Bakken, J.S.; Dumler, J.S. Clinical diagnosis and treatment of human granulocytotropic anaplasmosis. Ann. N. Y. Acad. Sci. 2006, 1078, 236–247. [Google Scholar] [CrossRef]
- Bakken, J.S.; Dumler, J.S. Human granulocytic anaplasmosis. Infect. Dis. Clin. 2015, 29, 341–355. [Google Scholar] [CrossRef]
- Buller, R.S.; Arens, M.; Hmiel, S.P.; Paddock, C.D.; Sumner, J.W.; Rikihisa, Y.; Unver, A.; Gaudreault-Keener, M.; Manian, F.A.; Liddell, A.M.; et al. Ehrlichia ewingii, a Newly Recognized Agent of Human Ehrlichiosis. N. Engl. J. Med. 1999, 341, 148–155. [Google Scholar] [CrossRef]
- Chen, S.; Yu, X.; Popov, V.L.; Westerman, E.L.; Hamilton, F.G.; Walker, D.H. Genetic and Antigenic Diversity of Ehrlichia chaffeensis: Comparative Analysis of a Novel Human Strain from Oklahoma and Previously Isolated Strains. J. Infect. Dis. 1997, 175, 856–863. [Google Scholar] [CrossRef]
- Chapes, S.K.; Ganta, R.R. Defining the immune response to Ehrlichia species using murine models. Vet. Parasitol. 2008, 158, 344–359. [Google Scholar] [CrossRef] [PubMed]
- Jankovic, D.; Kullberg, M.C.; Hieny, S.; Caspar, P.; Collazo, C.M.; Sher, A. In the absence of IL-12, CD4+ T cell responses to intracellular pathogens fail to default to a Th2 pattern and are host protective in an IL-10−/− setting. Immunity 2002, 16, 429–439. [Google Scholar] [CrossRef] [PubMed]
- Kuriakose, J.A.; Zhang, X.; Luo, T.; McBride, J.W. Molecular basis of antibody mediated immunity against Ehrlichia chaffeensis involves species-specific linear epitopes in tandem repeat proteins. Microbes Infect. 2012, 14, 1054–1063. [Google Scholar] [CrossRef] [PubMed]
- Velayutham, T.S.; Kumar, S.; Zhang, X.; Kose, N.; Walker, D.H.; Winslow, G.; Crowe, J.E., Jr.; McBride, J.W. Ehrlichia chaffeensis Outer Membrane Protein 1-Specific Human Antibody-Mediated Immunity Is Defined by Intracellular TRIM21-Dependent Innate Immune Activation and Extracellular Neutralization. Infect. Immun. 2019, 87, 12. [Google Scholar] [CrossRef]
- Available online: https://www.uptodate.com/contents/treatment-of-lyme-disease (accessed on 1 September 2022).
- Verhaegh, D.; Joosten, L.A.; Oosting, M. The role of host immune cells and Borrelia burgdorferi antigens in the etiology of Lyme disease. Eur. Cytok. Netw. 2017, 28, 70–84. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.epicentro.iss.it/zecche/borreliosi (accessed on 1 September 2022).
- Steere, A.C.; Malawista, S.E.; Snydman, D.R.; Shope, R.E.; Andiman, W.A.; Ross, M.R.; Steele, F.M. Lyme Arthritis: An Epidemic of Oligoarticular Arthritis in Children and Adults in Three Connecticut Communities. Arthr. Rheumatol. Eur. J. Epidemiol. 1977, 20, 7–17. [Google Scholar] [CrossRef]
- Benach, J.L.; Bosler, E.M.; Hanrahan, J.P.; Coleman, J.L.; Habicht, G.S.; Bast, T.F.; Cameron, D.J.; Ziegler, J.L.; Barbour, A.G.; Burgdorfer, W.; et al. Spirochetes Isolated from the Blood of Two Patients with Lyme Disease. N. Engl. J. Med. 1983, 308, 740–742. [Google Scholar] [CrossRef]
- Burgdorfer, W.; Barbour, A.G.; Hayes, S.F.; Benach, J.L.; Grunwaldt, E.; Davis, J.P. Lyme Disease—A Tick-Borne Spirochetosis? Science 1982, 216, 1317–1319. [Google Scholar] [CrossRef]
- Rizzoli, A.; Hauffe, H.C.; Carpi, G.; Vourc’h, G.I.; Neteler, M.; Rosà, R. Lyme borreliosis in Europe. Eurosurveillance 2011, 16, 19906. [Google Scholar] [CrossRef]
- Vandekerckhove, O.; De Buck, E.; Van Wijngaerden, E. Lyme disease in Western Europe: An emerging problem? A systematic review. Acta Clin. Belg. 2019, 76, 244–252. [Google Scholar] [CrossRef]
- D’Amico, W.; De Merich, D.; Di Renzi, S.; D’Ovidio, M.C.; Martini, A.; Melis, P.; Tomao, P.; Vonesch, N. Dipartimento di Medicina, Epidemiologia, Igiene del Lavoro e Ambientale. Inail. 2018. Available online: https://dsv.units.it/sites/dsv.units.it/files/Zoonosi-trasmesse-da-zecche_documento%20INAIL_12_07_2018_conEC.pdf (accessed on 17 July 2022).
- Sykes, R.A.; Makiello, P. An estimate of Lyme borreliosis incidence in Western Europe. J. Public Health 2017, 39, 74–81. [Google Scholar]
- Beltrame, A.; Rodari, P.; Mauroner, L.; Zanella, F.; Moro, L.; Bertoli, G.; Da Re, F.; Russo, F.; Napoletano, G.; Silva, R. Emergence of Lyme borreliosis in the province of Verona, Northern Italy: Five-years of sentinel surveillance. Ticks Tick-borne Dis. 2020, 12, 101628. [Google Scholar] [CrossRef]
- Cook, M.J. Lyme borreliosis: A review of data on transmission time after tick attachment. Int. J. Gen. Med. 2014, 8, 1–8. [Google Scholar] [CrossRef]
- Piesman, J.; Gern, L. Lyme borreliosis in Europe and North America. Parasitology 2004, 129, S191–S220. [Google Scholar] [CrossRef] [PubMed]
- Stanek, G.; Strle, F. Lyme borreliosis–from tick bite to diagnosis and treatment. FEMS Microbiol. Rev. 2018, 42, 233–258. [Google Scholar] [CrossRef] [PubMed]
- Steere, A.C. Lyme Disease. N. Engl. J. Med. 2001, 345, 115–125. [Google Scholar] [CrossRef]
- Steere, A.C.; Dhar, A.; Hernandez, J.; Fischer, P.A.; Sikand, V.K.; Schoen, R.T.; Nowakowski, J.; McHugh, G.; Persing, D.H. Systemic symptoms without erythema migrans as the presenting picture of early Lyme disease. Am. J. Med. 2003, 114, 58–62. [Google Scholar] [CrossRef]
- Branda, J.A.; Steere, A.C. Laboratory diagnosis of Lyme borreliosis. Clin. Microbiol. Rev. 2021, 34, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Aguero-Rosenfeld, M.E.; Wang, G.; Schwartz, I.; Wormser, G.P. Diagnosis of Lyme Borreliosis. Clin. Microbiol. Rev. 2005, 18, 484–509. [Google Scholar] [CrossRef]
- Bradley, J.F.; Johnson, R.C.; Goodman, J.L. The Persistence of Spirochetal Nucleic Acids in Active Lyme Arthritis. Ann. Intern. Med. 1994, 120, 487–489. [Google Scholar] [CrossRef] [PubMed]
- Nocton, J.J.; Dressler, F.; Rutledge, B.J.; Rys, P.N.; Persing, D.H.; Steere, A.C. Detection of Borrelia burgdorferi DNA by Polymerase Chain Reaction in Synovial Fluid from Patients with Lyme Arthritis. N. Engl. J. Med. 1994, 330, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Sigal, L.H. The polymerase chain reaction assay for Borrelia burgdorferi in the diagnosis of Lyme disease. Ann. Int. Med. 1994, 120, 520–521. [Google Scholar] [CrossRef] [PubMed]
- Nocton, J.J.; Bloom, B.J.; Rutledge, B.J.; Persing, D.H.; Logigian, E.L.; Schmid, C.H.; Steere, A.C. Detection of Borrelia burgdorferi DNA by Polymerase Chain Reaction in Cerebrospinal Fluid in Lyme Neuroborreliosis. J. Infect. Dis. 1996, 174, 623–627. [Google Scholar] [CrossRef]
- Guérin, M.; Shawky, M.; Zedan, A.; Octave, S.; Avalle, B.; Maffucci, I.; Padiolleau-Lefèvre, S. Lyme borreliosis diagnosis: State of the art of improvements and innovations. BMC Microbiol. 2023, 23, 204. [Google Scholar] [CrossRef]
- Callister, S.M.; Jobe, D.A.; Stuparic-Stancic, A.; Miyamasu, M.; Boyle, J.; Dattwyler, R.J.; Arnaboldi, P.M. Detection of IFN-γ secretion by T cells collected before and after successful treatment of early Lyme disease. Clin. Infect. Dis. 2016, 62, 1235–1241. [Google Scholar] [CrossRef] [PubMed]
- Lintner, H.; Hochgatterer-Rechberger, P.; Pischinger, B.; Seier, J.; Vollmann, P.; Haushofer, A.; Topakian, R. Sensitivity and specificity of cerebrospinal fluid CXCL13 for diagnosing Lyme neuroborreliosis-a study on 1410 patients and review of the literature. J. Neurol. Sci. 2020, 414, 116843. [Google Scholar] [CrossRef]
- Pilz, G.; Steger, R.; Wipfler, P.; Otto, F.; Afazel, S.; Haschke-Becher, E.; Harrer, A. Beyond LNB: Real life data on occurrence and extent of CSF CXCL13 in neuroinflammatory diseases. J. Neuroimmunol. 2020, 338, 577087. [Google Scholar] [CrossRef]
- Molins, C.R.; Ashton, L.V.; Wormser, G.P.; Hess, A.M.; Delorey, M.J.; Mahapatra, S.; Belisle, J.T. Development of a metabolic biosignature for detection of early Lyme disease. Clin. Infect. Dis. 2015, 60, 1767–1775. [Google Scholar] [CrossRef]
- Branda, J.A.; Lemieux, J.E.; Blair, L.; Ahmed, A.A.; Hong, D.K.; Bercovici, S.; Pollock, N.R. Detection of Borrelia burgdorferi cell-free DNA in human plasma samples for improved diagnosis of early Lyme borreliosis. Clin. Infect. Dis. 2021, 73, e2355–e2361. [Google Scholar] [CrossRef]
- Van de Schoor, F.R.; Baarsma, M.E.; Gauw, S.A.; Kullberg, B.J.; van den Wijngaard, C.C.; Hovius, J.W. Validation of cellular tests for Lyme borreliosis (VICTORY) study. BMC Infect. Dis. 2019, 19, 732. [Google Scholar] [CrossRef] [PubMed]
- Samuels, J.D.R.; Scott, D. Lyme Disease and Relapsing Fever Spirochetes: Genomics, Molecular Biology, Host Interactions and Disease Pathogenesis; Caister Academic Press: Wymondham, UK, 2021. [Google Scholar]
- Wormser, G.P.; McKenna, D.; Carlin, J.; Nadelman, R.B.; Cavaliere, L.F.; Holmgren, D.; Byrne, D.W.; Nowakowski, J. Brief Communication: Hematogenous Dissemination in Early Lyme Disease. Ann. Intern. Med. 2005, 142, 751–755. [Google Scholar] [CrossRef] [PubMed]
- Robin, J.; Bluth, M.; Ruditzky, M.; Norowitz, K.; Chice, S.; Pytlak, E.; Nowakowski, M.; Durkin, H.; Smith-Norowitz, T. IgE anti-Borrelia Burgdorferi components (p18, p31, p34, p41, p45, p60) and increased blood CD8+CD60+ T cells in children with lyme disease. J. Allergy Clin. Immunol. 2005, 115, S256. [Google Scholar] [CrossRef]
- Gomes-Solecki, M.; Arnaboldi, P.M.; Backenson, P.B.; Benach, J.L.; Cooper, C.L.; Dattwyler, R.J.; Schutzer, S.E.; Marconi, R.T.; Marques, A.R.; Molloy, P.; et al. Protective immunity and new vaccines for Lyme disease. Clin. Infect. Dis. 2020, 70, 1768–1773. [Google Scholar] [CrossRef] [PubMed]
- Gomes-Solecki, M. Blocking pathogen transmission at the source: Reservoir targeted OspA-based vaccines against Borrelia burgdorferi. Front. Cell. Infect. Microbiol. 2014, 4, 136. [Google Scholar] [CrossRef] [PubMed]
- Klouwens, M.J.; Trentelman, J.J.A.; Wagemakers, A.; Ersoz, J.I.; Bins, A.D.; Hovius, J.W. Tick-Tattoo: DNA Vaccination against B. burgdorferi or Ixodes scapularis Tick Proteins. Front. Immunol. 2021, 12, 615011. [Google Scholar] [CrossRef]
- Kaye, P.; Scott, P. Leishmaniasis: Complexity at the host–pathogen interface. Nat. Rev. Genet. 2011, 9, 604–615. [Google Scholar] [CrossRef]
- Available online: https://www.cdc.gov/parasites/leishmaniasis/index.html (accessed on 18 August 2022).
- Available online: https://www.salute.gov.it/portale/sanitaAnimale/dettaglioContenutiSanitaAnimale.jsp?lingua=italiano&id=220&tab=1 (accessed on 3 September 2022).
- Available online: https://www.izsvenezie.it/temi/malattie-patogeni/leishmaniosi/ (accessed on 3 September 2022).
- Cecílio, P.; Cordeiro-Da-Silva, A.; Oliveira, F. Sand flies: Basic information on the vectors of leishmaniasis and their interactions with Leishmania parasites. Commun. Biol. 2022, 5, 305. [Google Scholar] [CrossRef]
- World Health Organization. Leishmaniasis. Available online: https://www.who.int/health-topics/leishmaniasis#tab=tab_1 (accessed on 3 September 2022).
- Available online: https://www.epicentro.iss.it/leishmaniosi/epidemiologia (accessed on 3 September 2022).
- Available online: https://www.salute.gov.it/portale/sanitaAnimale/dettaglioContenutiSanitaAnimale.jsp?lingua=italiano&id=220&tab=2 (accessed on 18 August 2022).
- Available online: https://www.who.int/news/item/08-06-2022-visceral-leishmaniasis-and-HIV-coinfection-WHO-publishes-new-guideline-with-region-specific-treatment-recommendations (accessed on 13 September 2022).
- Salman, S.M.; Rubeiz, N.G.; Kibbi, A.-G. Cutaneous leishmaniasis: Clinical features and diagnosis. Clin. Dermatol. 1999, 17, 291–296. [Google Scholar] [CrossRef]
- Srivastava, P.; Dayama, A.; Mehrotra, S.; Sundar, S. Diagnosis of visceral leishmaniasis. Transact. R. Soc. Trop. Med. Hyg. 2011, 105, 1–6. [Google Scholar] [CrossRef]
- Saporito, L.; Giammanco, G.M.; De Grazia, S.; Colomba, C. Visceral leishmaniasis: Host–parasite interactions and clinical presentation in the immunocompetent and in the immunocompromised host. Int. J. Infect. Dis. 2013, 17, e572–e576. [Google Scholar] [CrossRef] [PubMed]
- Van Griensven, J.; Diro, E. Visceral leishmaniasis. Infect. Dis. Clin. 2012, 26, 309–322. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.E.; Streit, J.A. Visceral leishmaniasis. Gastroenterol. Clin. 1996, 25, 535–551. [Google Scholar] [CrossRef]
- Torres-Guerrero, E.; Quintanilla-Cedillo, M.R.; Ruiz-Esmenjaud, J.; Arenas, R. Leishmaniasis: A review. F1000Research 2017, 6, 750. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, J.N.; Lockwood, D.N. Clinical aspects of visceral leishmaniasis in HIV infection. Curr. Opin. Infect. Dis. 2013, 26, 1–9. [Google Scholar] [CrossRef]
- Abadías-Granado, I.; Diago, A.; Cerro, P.A.; Palma-Ruiz, A.M.; Gilaberte, Y. Leishmaniasis cutánea y mucocutánea. Actas Dermo-Sifiliográfic. 2021, 112, 601–618. [Google Scholar] [CrossRef]
- Dowlati, Y. Cutaneous leishmaniasis: Clinical aspect. Clin. Dermatol. 1996, 14, 425–431. [Google Scholar] [CrossRef]
- Kubba, R.; Al-Gindan, Y.; El-Hassan, A.; Omer, A. Clinical diagnosis of cutaneous leishmaniasis (oriental sore). J. Am. Acad. Dermatol. 1987, 16, 1183–1189. [Google Scholar] [CrossRef]
- Ahluwalia, S. Mucocutaneous leishmaniasis: An imported infection among travellers to central and South America. BMJ 2004, 329, 842–844. [Google Scholar] [CrossRef]
- Aronson, N.; Herwaldt, B.L.; Libman, M.; Pearson, R.; Lopez-Velez, R.; Weina, P.; Magill, A. Diagnosis and treatment of leishmaniasis: Clinical practice guidelines by the Infectious Diseases Society of America (IDSA) and the American Society of Tropical Medicine and Hygiene (ASTMH). Clin. Infect. Dis. 2016, 63, e202–e264. [Google Scholar] [CrossRef]
- Boggild, A.K.; Ramos, A.P.; Espinosa, D.; Valencia, B.M.; Veland, N.; Miranda-Verastegui, C.; Llanos-Cuentas, A. Clinical and demographic stratification of test performance: A pooled analysis of five laboratory diagnostic methods for American cutaneous leishmaniasis. Am. J. Trop. Med. Hyg. 2010, 83, 345. [Google Scholar] [CrossRef]
- Reithinger, R.; Dujardin, J.C. Molecular diagnosis of leishmaniasis: Current status and future applications. J. Clin. Microbiol. 2007, 45, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Sundar, S.; Rai, M. Laboratory diagnosis of visceral leishmaniasis. Clin. Vaccine Immunol. 2002, 9, 951–958. [Google Scholar] [CrossRef] [PubMed]
- Alvar, J.; Aparicio, P.; Aseffa, A.; Den Boer, M.; Canavate, C.; Dedet, J.P.; Moreno, J. The relationship between leishmaniasis and AIDS: The second 10 years. Clin. Microbiol. Rev. 2008, 21, 334–359. [Google Scholar] [PubMed]
- van Griensven, J.; Zijlstra, E.E.; Hailu, A. Visceral Leishmaniasis and HIV Coinfection: Time for Concerted Action. PLOS Neglected Trop. Dis. 2014, 8, e3023. [Google Scholar] [CrossRef]
- Burki, T. Guidelines for visceral leishmaniasis and HIV co-infection. Lancet Infect. Dis. 2022, 22, 1124–1125. [Google Scholar] [CrossRef]
- Dayakar, A.; Chandrasekaran, S.; Kuchipudi, S.V.; Kalangi, S.K. Cytokines: Key determinants of resistance or disease progression in visceral leishmaniasis: Opportunities for novel diagnostics and immunotherapy. Front. Immunol. 2019, 10, 670. [Google Scholar]
- Dos Santos, J.C.; de Figueiredo, A.M.B.; Silva, M.V.T.; Cirovic, B.; de Bree, L.C.J.; Damen, M.S.; Joosten, L.A. β-Glucan-induced trained immunity protects against Leishmania braziliensis infection: A crucial role for IL-32. Cell Rep. 2019, 28, 2659–2672. [Google Scholar] [CrossRef]
- Gueirard, P.; Laplante, A.; Rondeau, C.; Milon, G.; Desjardins, M. Trafficking of Leishmania donovani promastigotes in non-lytic compartments in neutrophils enables the subsequent transfer of parasites to macrophages. Cell. Microbiol. 2008, 10, 100–111. [Google Scholar] [CrossRef]
- Laufs, H.; Müller, K.; Fleischer, J.; Reiling, N.; Jahnke, N.; Jensenius, J.C.; Laskay, T. Intracellular survival of Leishmania major in neutrophil granulocytes after uptake in the absence of heat-labile serum factors. Infect. Immun. 2002, 70, 826–835. [Google Scholar] [CrossRef]
- Hawn, T.R.; Ozinsky, A.; Underhill, D.M.; Buckner, F.S.; Akira, S.; Aderem, A. Leishmania major activates IL-1α expression in macrophages through a MyD88-dependent pathway. Microbes Infect. 2002, 4, 763–771. [Google Scholar] [CrossRef] [PubMed]
- Matera, G.; Torti, C.; Mazzitelli, M.; Greco, G.; Rania, A.; Peronace, C.; Settembre, P.; Galati, L.; Giancotti, A.; Lamberti, A.G.; et al. Depression of lymphocyte activity during cutaneous leishmaniasis: A case report. Diagn. Microbiol. Infect. Dis. 2018, 92, 230–234. [Google Scholar] [CrossRef] [PubMed]
- Liese, J.; Schleicher, U.; Bogdan, C. The innate immune response against Leishmania parasites. Immunobiology 2008, 213, 377–387. [Google Scholar] [CrossRef]
- Machado, G.U.; Prates, F.V.; Machado, P.R.L. Disseminated leishmaniasis: Clinical, pathogenic, and therapeutic aspects. An. Bras. Dermatol. 2019, 94, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Fernández, L.; Carrillo, E.; Sánchez-Sampedro, L.; Sánchez, C.; Ibarra-Meneses, A.V.; Jimenez, M.A.; Moreno, J. Antigenicity of leishmania-activated C-kinase antigen (LACK) in human peripheral blood mononuclear cells, and protective effect of prime-boost vaccination with pCI-neo-LACK plus attenuated LACK-expressing Vaccinia viruses in hamsters. Front. Immunol. 2018, 9, 843. [Google Scholar]
- Krassowski, M.; Das, V.; Sahu, S.K.; Misra, B.B. State of the Field in Multi-Omics Research: From Computational Needs to Data Mining and Sharing. Front. Genet. 2020, 11, 610798. [Google Scholar] [CrossRef]
- Cuypers, B.; Meysman, P.; Erb, I.; Bittremieux, W.; Valkenborg, D.; Baggerman, G.; Mertens, I.; Sundar, S.; Khanal, B.; Notredame, C.; et al. Four layer multi-omics reveals molecular responses to aneuploidy in Leishmania. PLOS Pathog. 2022, 18, e1010848. [Google Scholar] [CrossRef]
- Li, R.; Ma, Z.; Zheng, W.; Wang, Z.; Yi, J.; Xiao, Y.; Chen, C. Multiomics analyses reveals Anaplasma phagocytophilum Ats-1 induces anti-apoptosis and energy metabolism by upregulating the respiratory chain-mPTP axis in eukaryotic mitochondria. BMC Microbiol. 2022, 22, 271. [Google Scholar]
- Vilanova, C.; Porcar, M. Are multi-omics enough? Nat. Microbiol. 2016, 1, 16101. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Epidemiological Criteria | Points |
|---|---|
| Staying in an endemic area | 2 |
| Occurrence between May and September | 2 |
| Contact with dog ticks | 2 |
| Clinical criteria | |
| Fever > 39 °C | 5 |
| Eschar | 5 |
| Maculo-papular or purpuric rash | 5 |
| Two of the three clinical criteria | 3 |
| All three clinical criteria present | 5 |
| Non-specific laboratory findings | |
| Platelets < 150,000/mm3 | 1 |
| ALT or AST > 50 u/L | 1 |
| Bacteriological criteria | |
| Isolation’s blood of R. conorii | 25 |
| R. conorii on immunofluorescence skin biopsy | 25 |
| Serological criteria | |
| Sole serum sample total Ig ≥ 1:128 | 5 |
| Sole serum sample IgM ≥ 1:160 and IgG ≥ 1:128 | 10 |
| Four-fold increase in paired serum samples | 20 |
| Type of Complications | Complication | Comment |
|---|---|---|
| Skeletal | Arthritis Spondylitis Sacroiliitis Osteomyelitis Bursitis Tenosynovitis | Occur in approximately 20–85% of patients. In children, arthritis of the hip and knee joints is most common. Unilateral sacroiliitis is common in young adults. Spondylitis is the most serious complication and paraspinal and epidural abscess are not infrequent. |
| Neuro-psychological | Meningoencephalitis Cerebral abscess Myelitis neuritis Depression Psychosis | Occur in approximately 2–5% of patients. Cerebrospinal fluid examination reveals lymphocytic pleocytosis with elevated protein and normal or low glucose level. Gram’s stain and culture are of low sensitivity. Computerized tomography may demonstrate basal ganglia calcification and abscesses. |
| Genitourinary | Epididymo-orchitis Prostatitis and cystitis Interstitial nephritis Glomerulonephritis | Unilateral epididymo-orchitis is frequent in young men. |
| Cardiovascular | Endocarditis (aortic > mitral) Myocarditis and pericarditis Endarteritis | Occurs in <2% of cases, but is the most common cause of death. Embolic phenomena are common. Valve replacement is warranted in most cases. Mycotic aneurisms of the aorta and large vessels are rare. |
| Hepatobiliary | Non-granulomatous and granulomatous hepatitis Abscesses Cirrhosis Acute cholecystitis | Abnormal liver function tests occur in 30–90% of patients. Percutaneous drainage and prolonged course of antibiotics. |
| Spleen | Splenomegaly Spleen abscess Splenic calcifications | Surgical drainage of localized suppurative lesions and splenectomy may be of value if antimicrobial treatment is ineffective. |
| Pulmonary | Hilar adenopathy Perihilar infiltrates Nodular lesions Lung abscess Interstitial pattern and pleural effusions | Cough and other pulmonary symptoms in approximately 15–25% of patients. |
| Hematological | Anemia Leucopenia Thrombocytopenia and pancytopenia Disseminated intravascular coagulation | More common in patients with Brucella melitensis. |
| Cutaneous | Rashes Papules Petechiae Purpura Cutaneous granulomatous vasculitis and erythema nodosum | Occur in about 5% of patients. Many transient and often non-specific skin lesions have been described. |
| Others | Uveitis and thyroiditis |
| Bartonella Factor | Function | Pathogenesis |
|---|---|---|
| TAAS (BadA, Vomps) | Host cell binding, attachment to extracellular matrix, activation of HIF-1, auto-aggregation, activation of NF-κB pathway, and phagocytosis inhibition | Strong interaction with host cells favors the secretion of proangiogenic cytokines |
| VirB/D4 IV Bartonella effector proteins | Inhibition of apoptosis mechanism, proinflammatory and angiogenesis activation | Control of the intracellular defense mechanism |
| OMP | NF-κB pathway activation | Overexpression of E-selectin and ICAM-1 factors may be involved in proangiogenic pathway activation |
| Deformin and Trw type IV secretion system | Adhesion and membrane invaginations of erythrocytes | Erythrocyte invasion |
| LPS | Antagonizes the TLR4 pathway | Immunomodulation |
| Symptom, Sign or Laboratory Finding | HME | HGA |
|---|---|---|
| % (n), Pts | % (n), Pts | |
| Symptom or sign | ||
| Fever | 97 (633) | 93 (521) |
| Myalgia | 57 (250) | 77 (516) |
| Headache | 80 (240) | 76 (385) |
| Malaise | 82 (234) | 94 (288) |
| Nausea | 64 (143) | 38 (258) |
| Vomiting | 33 (192) | 26 (901) |
| Diarrhea | 23 (197) | 16 (951) |
| Cough | 26 (155) | 19 (260) |
| Arthralgias | 41 (211) | 46 (504) |
| Rash | 31 (286) | 6 (357) |
| Stiff neck | 3 (240) | 21 (24) |
| Confusion | 19 (279) | 17 (211) |
| Laboratory finding | ||
| Leukopenia | 62 (276) | 49 (336) |
| Thrombocytopenia | 71 (247) | 71 (336) |
| Elevated serum AST or ALT level | 83 (276) | 71 (177) |
| Clinical Complication and Outcome | HME % Pts or Reports | HGA % Pts or Reports |
|---|---|---|
| Hospitalization | 42 | 33–50 |
| Intensive care unit admission | ND | 7 |
| Life-threatening complication | 17 | 7 |
| Death | 3 | <1 |
| Hemodynamic system complication Toxic or septic-shock syndrome Coagulopathy Hemorrhage Myocarditis or heart failure Renal failure | ++ 2 + + 2 | + <1 ± ± 1 |
| Pneumonia ARDS | 1 | 1 |
| CNS and PNS Complications Meningoencephalitis Cranial nerve palsies Demyelinating polyneuropathy Brachial plexopathy | 4 ± NR NR | 1 ± ± ± |
| Acute abdominal syndrome | + | NR |
| Rhabdomyolysis | NR | ± |
| Opportunistic infection | + | ++ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lionello, F.C.P.; Rotundo, S.; Bruno, G.; Marino, G.; Morrone, H.L.; Fusco, P.; Costa, C.; Russo, A.; Trecarichi, E.M.; Beltrame, A.; et al. Touching Base with Some Mediterranean Diseases of Interest from Paradigmatic Cases at the “Magna Graecia” University Unit of Infectious Diseases: A Didascalic Review. Diagnostics 2023, 13, 2832. https://doi.org/10.3390/diagnostics13172832
Lionello FCP, Rotundo S, Bruno G, Marino G, Morrone HL, Fusco P, Costa C, Russo A, Trecarichi EM, Beltrame A, et al. Touching Base with Some Mediterranean Diseases of Interest from Paradigmatic Cases at the “Magna Graecia” University Unit of Infectious Diseases: A Didascalic Review. Diagnostics. 2023; 13(17):2832. https://doi.org/10.3390/diagnostics13172832
Chicago/Turabian StyleLionello, Ferdinando Carmelo Pio, Salvatore Rotundo, Gabriele Bruno, Gabriella Marino, Helen Linda Morrone, Paolo Fusco, Chiara Costa, Alessandro Russo, Enrico Maria Trecarichi, Anna Beltrame, and et al. 2023. "Touching Base with Some Mediterranean Diseases of Interest from Paradigmatic Cases at the “Magna Graecia” University Unit of Infectious Diseases: A Didascalic Review" Diagnostics 13, no. 17: 2832. https://doi.org/10.3390/diagnostics13172832